 + Acanthostega
+ Acanthostega  .
.| Tetrapoda | ||
| The Vertebrates | Overview |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Sarcopterygii
│
└─Tetrapoda
├─Ichthyostega
└─┬─Tulerpeton
└─┬─Crassigyrinus
└─┬─Colosteidae
└─┬─Baphetidae
└─Tetrapoda*
├─Temnospondyli
└─┬─Lepospondyli
└─Reptiliomorpha
|
Contents
Overview |
| Sarcopterygii
├─Lungfishes
└─Tetrapoda #1
├─Rhizodontiformes
└─┬─Tristichopteridae
│ ├─Eusthenopteron
│ └─Mandageria
└─┬─Panderichthys
└─Tetrapoda #2
├─Livoniana
└─┬─Elginerpeton
└─┬─Obruchevichthys
└─Tetrapoda #3 & Tetrapoda
├─Acanthostega
└─Tetrapoda #4 & Tetrapoda*
├─Temnospondyls
│ ├─salamanders
│ └─frogs
└─┬─snakes
└─Pander, princes, professors, etc. |
Abbreviated cladogram showing relative position of taxa |
The short answer is that we don't know. There are at least four definitions current in the scientific literature. These are from most to least inclusive):
1) Lizards > Lungfish. This is the "total group" definition, which aims to include the crown group and the entire stem. Coates et al. (2002). This definition has two major problems. First, this taxon already has a perfectly good name: Tetrapodomorpha. Second, it's a bit awkward to call something that is obviously a fish and has no feet -- Eusthenopteron for example -- a "tetrapod." Worse, this definition includes animals such as the more derived rhizodonts and tristichopterids. These forms scarcely have four paired fins, much less four feet.
2) Christian Pander > Panderichthys. This is a slightly obsolete attempt at a compromise position. We like it, except for (as usual) two reasons. First, Panderichthys is a very tetrapod-like fish. However, the definition assumes that Panderichthys is more or less on the direct line to conventional tetrapods. This is quite likely, but not quite as likely as it seemed a few years ago. Second, there is a growing gap between Panderichthys and the folks with four feet. Paleontologists are all constitutionally incapable of leaving a good thing alone. Now we have Livoniana, Elginerpeton, Obruchevichthys, and whatever comes out of the Baltics next year, to somehow cram in between Panderichthys and "conventional" tetrapods.
3) Animals with four feet and other characters. This is the historical or conventional definition. It suffers from the same problem as all apomorphy-based definitions. What if feet evolved more than once? When, exactly, does a fin become a foot? What about snakes or microsaurs? Finally, it is an arbitrary box. See discussion at Dendrograms.
4) frogs + princes. This is the crown group Tetrapoda, the last common ancestor of Lissamphibia and Amniota and all of its descendants. We like this one, too, but it's borders are extremely unclear, and it probably leaves far too many "labyrinthodont" forms homeless. As a practical matter, this definition is unuseful because it really irritates certain people, notably, Dr. Jenny Clack Similarly it seems to inspire an equal measure of tenacious defense from others, notably Prof. Clack's former coworker in the study of Acanthostega, Prof. Michael Coates [1].
So, in a really pathetic and sycophantic attempt to please everyone, this is what we're going to do:
+ Acanthostega . + frogs
+ frogs  .
.In Tetrapoda*, the asterisk is supposed to suggest a crown, since this is the crown group. Of course it suggests nothing of the sort, but life is full of these little disappointments. If one prefers, Tetrapoda* can be pronounced "crown tetrapods." Under this regime, the unornamented Tetrapoda is more or less the same as the colloquial "tetrapod." However, if we ever find the feet/fins of Livoniana and it has toes, that will be just too bad. It still won't be a tetrapod. If one prefers, Tetrapoda can be pronounced "stem tetrapods." ATW030921.
 Note added in disproof: since the original posting of this essay, Prof. Coates has decamped from his previous position on the issue, slipping away without so much as a postcard to Palaeos. Doubtless he hoped to be relieved of the onerous responsibilities of anchoring the Crown Tetrapoda*. Prof. Coates now seems to favor the "total group" concept ("Tetrapoda #1"). Ruta et al. (2003). We are unsure precisely where Prof. Clack currently stands, but she still stands on someone's feet. That is, she yet maintains that tetrapods may be defined as sarcopterygians with hands and feet. Clack 2002). However, the ground has shifted under those feet a bit, since it is now quite likely that, for example, Ventastega and Elginerpeton would qualify as tetrapods by this definition. Thus, she has been moved a few feet down the phylogenetic tree. Nonetheless, while our anchor taxa may have have abandoned their posts and turned their backs on their plain phylogenetic duty, we are unmoved. Like Horatius Cocles at the Pons Sublicius, we will stand our ground until the whole phylogenetic structure gives way and we, with our harness on our back, plunge headlong in the tide [2]. ATW050520.
Note added in disproof: since the original posting of this essay, Prof. Coates has decamped from his previous position on the issue, slipping away without so much as a postcard to Palaeos. Doubtless he hoped to be relieved of the onerous responsibilities of anchoring the Crown Tetrapoda*. Prof. Coates now seems to favor the "total group" concept ("Tetrapoda #1"). Ruta et al. (2003). We are unsure precisely where Prof. Clack currently stands, but she still stands on someone's feet. That is, she yet maintains that tetrapods may be defined as sarcopterygians with hands and feet. Clack 2002). However, the ground has shifted under those feet a bit, since it is now quite likely that, for example, Ventastega and Elginerpeton would qualify as tetrapods by this definition. Thus, she has been moved a few feet down the phylogenetic tree. Nonetheless, while our anchor taxa may have have abandoned their posts and turned their backs on their plain phylogenetic duty, we are unmoved. Like Horatius Cocles at the Pons Sublicius, we will stand our ground until the whole phylogenetic structure gives way and we, with our harness on our back, plunge headlong in the tide [2]. ATW050520.
[1] and our own colleague, the normally steady and convivial Mikko Haaramo, who has strongly protested our former use of "Tetrapodomorpha" to mean four-footed stem tetrapods.
[2] Lord Macauley's Horatius is only average poetry, but still a hell of a story.
 We usually leave this sort of anatomical survey for the "Characters" section of the main entries. However, so many stray bits and pieces have been added to our knowledge of early tetrapods over the last 15 years that we thought it might be interesting and convenient to sort them out here. That is, of course, interesting and convenient for us. We reluctantly concede that it may not be interesting (much less convenient) for the reader. Fortunately, one of the compensations of Palaeos is that, like a small chorus of demented bullfrogs shrouded in the fetid mists of some starless midnight bog, we enjoy a certain freedom to emit loud, coarse noises for the manifest satisfaction of doing so. Such is our response to the conjectural disapprobation of our equally conjectural readers.
We usually leave this sort of anatomical survey for the "Characters" section of the main entries. However, so many stray bits and pieces have been added to our knowledge of early tetrapods over the last 15 years that we thought it might be interesting and convenient to sort them out here. That is, of course, interesting and convenient for us. We reluctantly concede that it may not be interesting (much less convenient) for the reader. Fortunately, one of the compensations of Palaeos is that, like a small chorus of demented bullfrogs shrouded in the fetid mists of some starless midnight bog, we enjoy a certain freedom to emit loud, coarse noises for the manifest satisfaction of doing so. Such is our response to the conjectural disapprobation of our equally conjectural readers.
Having disposed of all opposition with the irrefutable logic of this batrachian simile, we will turn to the question of the day: why is Prof. Clack like Acanthostega? Or, to be more precise, what did their last common ancestor look like? To address this matter, we need no longer resort to batrachian simile. It is no longer necessary to invent an ancestor based on some ill-conceived analogy to frogs. The growing legion of Late Devonian 'stegas and 'petons -- particularly Acanthostega -- has yielded enough data that we can be quite specific.
In order to illustrate the tetrapod condition, we will need to make the close acquaintance of Auntie Stega, the last common ancestor of Acanthostega and ourselves. Auntie lived about 365 Mya, quite possibly in Baltica. We will suppose (for no particular reason) that she was Irish.
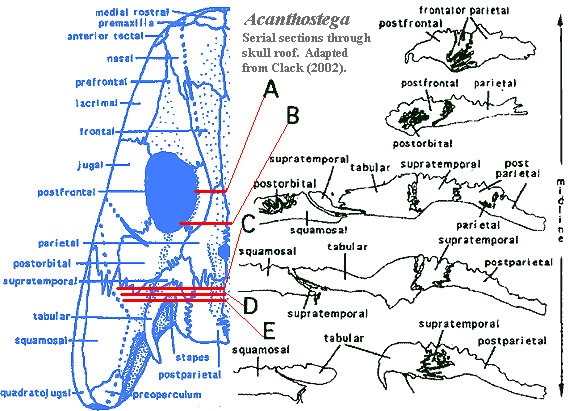 Auntie was a headstrong creature. That is, she had a heavily armored skull. The dermal bones locked together through complex, interdigitating sutures [C02a]. The word "interdigitating" doesn't really do justice to these structures. The word brings to mind interlocked fingers. This is a fair description of the skull sutures in many living amniotes, but it understates the weirdly complex, intertwined curves of Paleozoic forms. Auntie Stega thus surely had a hard head, but was somewhat inflexible by today's standards. In addition to being rigid, Auntie Stega was probably somewhat thin-skinned. Auntie had already evolved the characteristic tetrapod ornament of irregular pits and ridges, radiating from presumed growth centers [LC93] [A95]. Then again, she was perhaps a bit less ornamented than her Carboniferous descendants, which suggests a thicker and/or looser skin covering. However, the whole issue of skin covering is speculative, and many reconstructions seem to assume an absence of integument on the skull.
Auntie was a headstrong creature. That is, she had a heavily armored skull. The dermal bones locked together through complex, interdigitating sutures [C02a]. The word "interdigitating" doesn't really do justice to these structures. The word brings to mind interlocked fingers. This is a fair description of the skull sutures in many living amniotes, but it understates the weirdly complex, intertwined curves of Paleozoic forms. Auntie Stega thus surely had a hard head, but was somewhat inflexible by today's standards. In addition to being rigid, Auntie Stega was probably somewhat thin-skinned. Auntie had already evolved the characteristic tetrapod ornament of irregular pits and ridges, radiating from presumed growth centers [LC93] [A95]. Then again, she was perhaps a bit less ornamented than her Carboniferous descendants, which suggests a thicker and/or looser skin covering. However, the whole issue of skin covering is speculative, and many reconstructions seem to assume an absence of integument on the skull.
Auntie Stega's snout was probably broad, certainly low, and relatively long [LC93]. Her external nostrils were set very low, near the edge of the upper jaw bones. Id. These bones were probably rather narrow. We may imagine her as viewing the approaching close of the Devonian with pursed lips, nostrils flaring slightly with stern disapproval.
Despite her very modest intellectual attainments, Auntie had a well-developed skull table, generally of the conventional tetrapod sort, with successive pairs of nasals, frontals, parietals and postparietals. At the anterior end, Auntie Stega may have had paired rostrals in addition to the usual premaxillae. The nasals were a simple paired set, elongate and rather large [Co96]. However, she had a soft spot between them, an internasal fontanelle [C02a] [AL04]. This region was not bone- covered, and may have served to relieve some of the stresses of Irish Devonian life on her otherwise rigid skull. She had a few more bones than the usual amniote might have on the top of her head, but nothing flashy. These were just the traditional Paleozoic complement of intertemporals, supratemporals and the like -- all of the old-fashioned bones that became obsolete when the skull table later separated from the cheek.
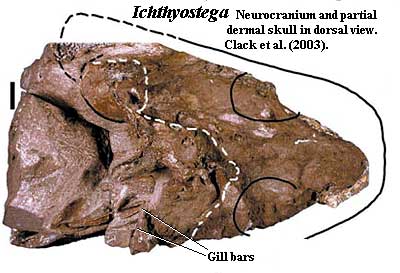
Auntie Stega's postorbital skull was markedly shorter than the skulls of her ancestors [C02a]. In part, this was due to the separation of the shoulder girdle from the skull and the reduction of the branchial (gill) region. This separation of head and shoulders may not have been complete in Auntie's day.She was a stiff-necked sort, at the very least. Nevertheless, the operculogular series was absent, with only a small preopercular remnant [Co96]. Thus, Auntie Stega could likely move her jaw without making an unseemly show of flapping her gill chambers or waving her arms. Yet she had functional internal gills, which she would use when she thought no one was looking. Indeed, her branchial apparatus included a large ceratohyal and three or more deeply grooved, ossified branchial arches [CoC91] [Co96] [C+03]. Quite likely she also had a functional spiracle, and may have sported a small tabular "horn" supporting the soft tissue of a spiracular notch [CoC91] [LC93].
 Auntie had large eyes, or, at least, her orbits were large. Likely, the orbits faced dorsally, although it isn't really possible to tell what her range of vision may have been. It is difficult to imagine Auntie Stega with great, adoring eyes cast ever heavenward, like a plaster saint. In addition to the practical difficulties of this pose, it seems entirely out of character.
Auntie had large eyes, or, at least, her orbits were large. Likely, the orbits faced dorsally, although it isn't really possible to tell what her range of vision may have been. It is difficult to imagine Auntie Stega with great, adoring eyes cast ever heavenward, like a plaster saint. In addition to the practical difficulties of this pose, it seems entirely out of character.
Not only was Auntie Stega's postorbital skull short, it was also much simplified. The postorbital and jugal were large and formed the entire posterior margin of the orbit [C02a] [A98]. The jugal also dominated the cheek region. Having finally established contact with the quadratojugal at the level of Ventastega, the jugal thereafter strictly forbade any contact between the squamosal and maxilla [Co96]. Thus the basic four-part organization of the tetrapod cheek was established: with postorbital and squamosal above, jugal and quadratojugal below.
We know remarkably little about Auntie Stega's occiput. As mentioned elsewhere, the formation of her neck was far from complete. The notochord still penetrated well into the skull [C98] and must yet have borne a considerable fraction of the task of holding up the head. We might assume that the occipital region looked like the occiput of Acanthostega. However, the posterior skull in the closely related Ichthyostega is so weirdly different [J96] [C+03] that such an assumption would be risky. Auntie Stega could not stick her neck out, and neither shall we.
 Auntie Stega's immediate ancestors were of two minds, with the sphenethmoid anterior) and otoccipital (posterior) regions separated by a fissure, the intracranial joint. Auntie had a more integrated personality. That is to say, her mind was made up, with the various parts of the braincase firmly sutured, or even fused, together. In particular, the sphenethmoid and the otic capsules were co-ossified [C98]. Except in Ichthyostega, the basisphenoid was sutured both to the basioccipital and to the prootic portion of the otic capsule [C98]. In turn, a prootic process braced the braincase on the parietal, and (except in Acanthostega) the opisthotic met a facet of the tabular [C98]. Auntie did not have one final piece of stapling that her descendants soon acquired, a parasphenoid which extended onto the otoccipital region [C94].
Auntie Stega's immediate ancestors were of two minds, with the sphenethmoid anterior) and otoccipital (posterior) regions separated by a fissure, the intracranial joint. Auntie had a more integrated personality. That is to say, her mind was made up, with the various parts of the braincase firmly sutured, or even fused, together. In particular, the sphenethmoid and the otic capsules were co-ossified [C98]. Except in Ichthyostega, the basisphenoid was sutured both to the basioccipital and to the prootic portion of the otic capsule [C98]. In turn, a prootic process braced the braincase on the parietal, and (except in Acanthostega) the opisthotic met a facet of the tabular [C98]. Auntie did not have one final piece of stapling that her descendants soon acquired, a parasphenoid which extended onto the otoccipital region [C94].
As we might expect from the short postorbital skull, the entire otic region was relatively short, front-to-back [C+03]. In fact, this region shows a remarkable degree of reorganization over a rather small region of phylospace [A+96]. The lateral commisure, otic shelf, and jugular canal -- standard features of Panderichthys and other osteolepiforms -- were all lost in Auntie's generation [C98]. The vestibular fontanelle was reorganized as a space that would eventually become the inner ear [Co96]. It opened to the outer world through what is presumed to be a fenestra ovalis. Despite all this talk of ears, Auntie Stega herself was probably quite deaf. The lateral otic fissure (embryonic metotic fissure of amniotes) was continuous with the inner ear, as it still is. However, the elaborate systems which allow amniotes to use this connection to sort and analyze sound waves did not yet exist [MC04].
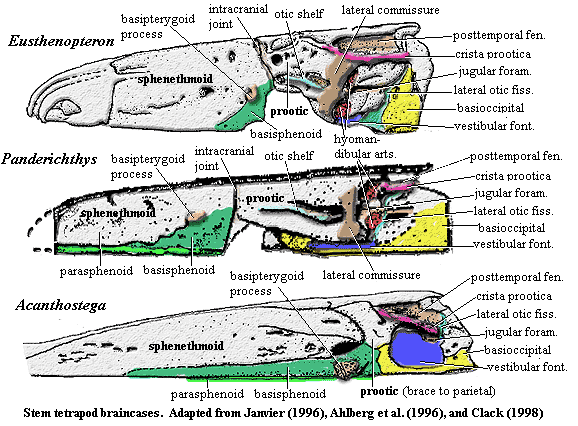 Perhaps because Auntie Stega was a single-minded sort, her mind had fewer facets. Specifically, she lacked the usual facets for the hyomandibula to attach to the braincase. Fortunately, she really didn't have much use for the hyomandibula anyway, since the whole branchial apparatus was of decreasing importance to her way of life. Neurocrania lack closets, umbrella stands, or other receptacles for elongate objects of no immediate utility, so the hyomandibula was more or less wadded up and thrown in a corner. One end came to rest blocking the vestibular fontanelle (or fenestra ovalis), while the distal end was jammed up against the tabular (or in that general vicinity), where it spread out, like a candle in holder which has fallen over and half melted on the table [C89]. This crude and unsightly condition is obviously a long way from the graceful double-headed arc of a proper hyomandibula. Perhaps for that reason, this bone is referred to, from Auntie's time onward, as the stapes.
Perhaps because Auntie Stega was a single-minded sort, her mind had fewer facets. Specifically, she lacked the usual facets for the hyomandibula to attach to the braincase. Fortunately, she really didn't have much use for the hyomandibula anyway, since the whole branchial apparatus was of decreasing importance to her way of life. Neurocrania lack closets, umbrella stands, or other receptacles for elongate objects of no immediate utility, so the hyomandibula was more or less wadded up and thrown in a corner. One end came to rest blocking the vestibular fontanelle (or fenestra ovalis), while the distal end was jammed up against the tabular (or in that general vicinity), where it spread out, like a candle in holder which has fallen over and half melted on the table [C89]. This crude and unsightly condition is obviously a long way from the graceful double-headed arc of a proper hyomandibula. Perhaps for that reason, this bone is referred to, from Auntie's time onward, as the stapes.
If the stapes didn't look much like a hyomandibula, it also bore little resemblance to the stapes of amniotes, among whom the stapes eventually became a tiny, gracile hearing ossicle for conducting sound from the tympanic membrane and through the middle ear. There is no indication that Auntie or any of her immediate descendants possessed a tympanic membrane [C89] [MC04]. Auntie Stega's stapes was thick, massive, and incapable of transmitting high frequency sound [C89] [MC04]. Perhaps the only resemblance to the future hearing ossicle was the stapedial foramen. Both heads of the old hyomandibula participated in blocking the fenestra ovalis, and the stapedial foramen appears to have been a relic of the former space between the two heads [C94]. Functionally, the stapes of Auntie's day was simply a mechanical brace between the braincase and the dermal skull.
Auntie Stega minded her mouth. Auntie's basipterygoid process was huge, bulbous, and anchored her braincase to her palate in an authoritative fashion by fitting into a socket formed by the epipterygoid and pterygoid [C98] [C02]. Originally, this may have been a moveable synovial joint [C89]. However, it was only used on holidays and eventually became an immoveable bolt in almost all of Auntie's descendants [LCo95]. The sphenethmoid region was already losing bone in Auntie Stega, and (to judge from Acanthostega) was also becoming quite thin and less massive. In all of Auntie's Carboniferous descendants, the sphenethmoid is largely unossified and thus unknown. Fortunately, Acanthostega retained enough bone in the sphenethmoid region to demonstrate that it was already acquiring the eye musculature of tetrapods, quite likely an adaptation to vision above the surface of the water.
Before biting into on the palate and jaw, we pause for some thoughts on the brain. What's going on here? The braincase has become suddenly much more compact, unitary and tightly braced. The front part is stripped to its sensory core, losing bone and mass. The relative length of the otic region is cut by about half. Two massive braces are added, and a line of weakness between the two halves is completely obliterated -- all in about 10 My and possibly much less. This is evolution on steroids. Like the skull bones -- but even more so -- the braincase has become somewhat simplified, tightly interlinked, and vastly stronger. We can think of only two factors that would be sufficient to explain this evolutionary sprint, and suggest that both were true. First, the skull was far more active because it was used in very active locomotion as the primary control surface, as argued elsewhere. Second, the skull was frequently uncushioned by water and was required to bear the more severe shocks of rapid momentum changes in air. [Recall -- if recollecting physics is not too painful an exercise -- that F = dp/dt. Force is rate change of momentum.] We don't mean to suggest that Auntie Stega was terrestrial; but she may well have kept her chin up and, perhaps aware of her status as the Mother of all Tetrapods, her nose in the air.
checked ATW031011