|
|
Bones: The Braincase |
| The Vertebrates |
The Occiput |
The Occiput
Anatomy
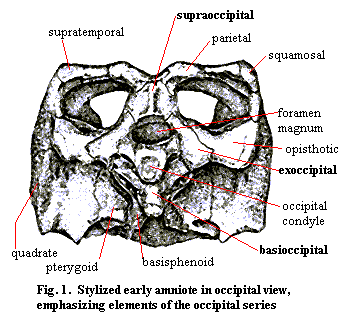 The occiput is most typically a series of four endochondral bones: the supraoccipital, the basioccipital and the paired exoccipitals. Together, or in various combinations, they usually form the occipital condyle(s), the cranial side of the articulation of the skull with the vertebral column, and define the boundaries of the foramen magnum (Latin for "big hole") where the spinal nerves exit the brain. Together with the quadrates, the opisthotics and various dermal bones, such as the parietals and the squamosals, the bones of the occiput form the back of the skull. A typical arrangement, that of our old friend, Bob the Basal Amniote, is shown in Figure 1. [1]
The occiput is most typically a series of four endochondral bones: the supraoccipital, the basioccipital and the paired exoccipitals. Together, or in various combinations, they usually form the occipital condyle(s), the cranial side of the articulation of the skull with the vertebral column, and define the boundaries of the foramen magnum (Latin for "big hole") where the spinal nerves exit the brain. Together with the quadrates, the opisthotics and various dermal bones, such as the parietals and the squamosals, the bones of the occiput form the back of the skull. A typical arrangement, that of our old friend, Bob the Basal Amniote, is shown in Figure 1. [1]
The triangular configuration of the three dorsal elements is fairly typical and also rather easy to understand. The supraoccipital ties the occiput to the dermal skull table, while the exoccipitals are braced, directly or through the opisthotics, to the quadrate, squamosal, and whatever other elements form the posterolateral columns of the skull. Together, the supraoccipital and the exoccipitals define the top and sides of the foramen magnum.
Bob's occipital condyle is formed entirely by the basioccipital. The exoccipitals also frequently participate with the basioccipital in the occipital condyle, particularly when there is more than one craniospinal articulation. Bob's condyle is also a rather simple socket for a basic ball-in-socket articulation. However, the rest of the basioccipital is a very complicated affair. The "pockets" flanking the condyle are probably shaped to accept elements of the first cervical vertebra which limit lateral and/or dorsoventral rotation of the skull with respect to the spinal column and resist hyperextension of the neck. These brakes on rotation are critical to prevent bending stresses on the spinal cord. In addition, the basioccipital must be braced strongly against compression by articulating with basicranial elements such as the basisphenoid and pterygoid. The design problem is acute, because the basioccipital is performing its structural duties dorsally, and there is no direct ventral support.
Functions
Most of the functions of the occiput have been discussed above:
1) formation of the foramen magnum. In lower vertebrates, the occiput or its functional equivalent) is organized around the penetration of the anterior notochord into the cranium, and separate foramina for the major vessels and the spinal nerve cord. However, the same principles apply.
2) formation of the craniospinal articulation, including the occipital condyle(s) and accessory articulations primarily designed to keep rotation, flexion, and compression of the skull with respect to the spine within a physiologically safe range.
3) structural support for the rear wall of the skull. The occiput usually has considerable help here from other elements.
4) structural support for the otic capsules, palate and basicranium as well as vice-versa).
Phylogeny
Chondrichthyes
 Although the standard occipital bones are a more recent phenomena, all craniates must, by definition, have an articulation between the braincase and the notochord (or vertebral column). Actually, it appears that the articulation in more basal vertebrates was far more complex than in Recent tetrapods. We are accustomed to think of the occipital articulation as involving two elements: (a) a mechanical coupling with the vertebrae and (b) a neural coupling with the spinal cord. In more basal forms, there were actually five different couplings which are (roughly from ventral to dorsal): (a) the dorsal aorta, (b) a cotylus or pit for the anterior notochord, (c) connections for the XIth cranial nerve the spinal accessory or occipitospinal nerve), (d) mechanical coupling for the vertebrae, and (e) the foramen magnum for the spinal cord. The occiput of the Lower Carboniferous shark Akmonistion, shown in Figure 2, illustrates one such arrangement. See Coates & Sequeira (1998). The very complexity of this arrangement may have created the need for an otico-occipital fissure to prevent excessive strain on the various components. Modern sharks are notably different. As shown in
Although the standard occipital bones are a more recent phenomena, all craniates must, by definition, have an articulation between the braincase and the notochord (or vertebral column). Actually, it appears that the articulation in more basal vertebrates was far more complex than in Recent tetrapods. We are accustomed to think of the occipital articulation as involving two elements: (a) a mechanical coupling with the vertebrae and (b) a neural coupling with the spinal cord. In more basal forms, there were actually five different couplings which are (roughly from ventral to dorsal): (a) the dorsal aorta, (b) a cotylus or pit for the anterior notochord, (c) connections for the XIth cranial nerve the spinal accessory or occipitospinal nerve), (d) mechanical coupling for the vertebrae, and (e) the foramen magnum for the spinal cord. The occiput of the Lower Carboniferous shark Akmonistion, shown in Figure 2, illustrates one such arrangement. See Coates & Sequeira (1998). The very complexity of this arrangement may have created the need for an otico-occipital fissure to prevent excessive strain on the various components. Modern sharks are notably different. As shown in 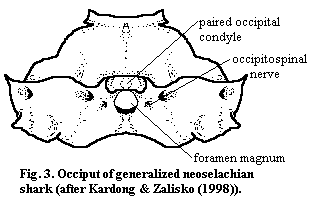 Figure
3, the notochord and dorsal aorta have been re-engineered so as not to participate directly in the occiput, and the occipital condyle has become a more prominent, paired structure dorsolateral to the foramen magnum.
Figure
3, the notochord and dorsal aorta have been re-engineered so as not to participate directly in the occiput, and the occipital condyle has become a more prominent, paired structure dorsolateral to the foramen magnum.
Osteichthyes
We have spent some extra time with the sharks in order to focus on the structure of the occiput as a whole, without the distraction of individual bones. The osteichthyans actually show a rather similar pattern of development, with mammals eventually arriving at an occipital structure that is somewhat similar to the neoselachian condition. Unfortunately (slipping, momentarily, out of the editorial "we") I have been unable to come up with enough information on the basic actinopterygian or sarcopterygian plans to comment much except to make the following tentative points. (1) The supraoccipital does not seem to have been part of the original complement of occipital ossifications. In teleosts, the supraoccipital is not clearly part of the occiput at all, but tends to form an anteroposterior crest on the top of the skull -- anchored in, but not part of, the occiput. The space between the foramen magnum and the roof of the skull (specifically, the post-parietals) is occupied by the opisthotic. 2) As in sharks, the dorsal aorta is lost from the occiput first, presumably because it is functionally replaced by the carotid arteries.
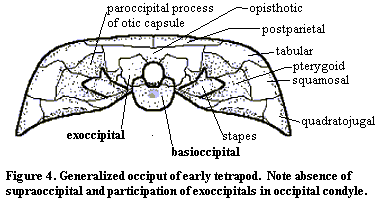 A generalized early tetrapod occiput is shown in Figure 4. Like the opercular bones, the notochordal cotyle of the occiput vanishes with or perhaps even before, the transition to land. In fact, it disappears almost as soon as there are any tetrapods to be found. Note that the exoccipitals participate in the occipital condyle -- a feature which is present, for example, in the eogyrinid Palaeoherpeton and becomes quite common in amniotes.
A generalized early tetrapod occiput is shown in Figure 4. Like the opercular bones, the notochordal cotyle of the occiput vanishes with or perhaps even before, the transition to land. In fact, it disappears almost as soon as there are any tetrapods to be found. Note that the exoccipitals participate in the occipital condyle -- a feature which is present, for example, in the eogyrinid Palaeoherpeton and becomes quite common in amniotes.
At least in the colosteid Greererpeton, the exoccipitals articulate directly with the postparietals, which may participate directly in the foramen magnum. In slightly more derived forms, the opisthotic appears on the surface of the occiput and separates the foramen magnum from the postparietals. In amniotes, the supraoccipital is added as well. This is consistent with the common observation that the height of the braincase tends to increase in more derived tetrapods*. Note that, although the global trend in tetrapods is to reduce the number of cranial bones, bones are actually being added in the occiput.
 The general pattern of the lepidosaur occiput is essentially the same as in early tetrapods, except that (a) the supraoccipital replaces the opisthotic and the opisthotic becomes a paired element lateral to the exoccipitals. In other words, the pattern is the same as for Bob the Basal Amniote.
The general pattern of the lepidosaur occiput is essentially the same as in early tetrapods, except that (a) the supraoccipital replaces the opisthotic and the opisthotic becomes a paired element lateral to the exoccipitals. In other words, the pattern is the same as for Bob the Basal Amniote.
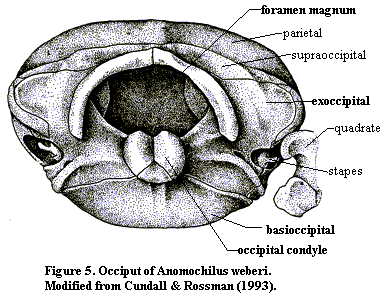
Figure 5 shows the same basic pattern in Anomochilus, a fairly basal snake (modified from Cundall & Rossman (1993)). As those authors note, "the posterior braincase of Anomochilus appears remarkable in shape but unremarkable in organization." Id. at 246. In more derived snakes, most of the occipital elements are fused, forming essentially the only akinetic structure in the skull. However, the organization of the occiput remains the same.
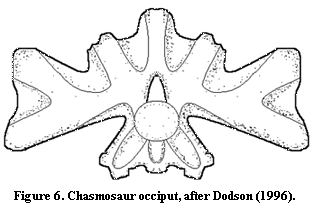 The archosaur occiput also differs very little from the basic tetrapod pattern. Frequently, the occipital elements are fused, so that the occiput appears to be a single block. In particular, the exoccipitals are usually completely fused with the opisthotics, always so in theropods. Currie & Zhao (1993). Consistent with that trend, there is a tendency for some foramina disappear so that, for example, the XIth nerve no longer exits the brain in the occiput itself. In addition, the occipital condyle tends to be in a more dorsal position relative to the skull as a whole. This does not appear to be due to restructuring of the occiput so much as to the elongation of the quadrate ventrally. That is, the skull becomes deeper while the occiput remains more or less unaffected. Finally, the fused exoccipital-opisthotic element forms a broad bar joining the occiput to the quadrates on either side. Because of the increasing fenestration of the back of the skull, the entire occipital complex may appear to form a sort of Maltese cross, as in the rough sketch of the occiput of Chasmosaurus in Figure
6 (after Dodson (1996)). Here, much of the edge of the occiput is not in contact with any other bone. The lateral edges articulate with the quadrates, the very top is attached to the parietals, and the base rests more broadly on the basisphenoid.
The archosaur occiput also differs very little from the basic tetrapod pattern. Frequently, the occipital elements are fused, so that the occiput appears to be a single block. In particular, the exoccipitals are usually completely fused with the opisthotics, always so in theropods. Currie & Zhao (1993). Consistent with that trend, there is a tendency for some foramina disappear so that, for example, the XIth nerve no longer exits the brain in the occiput itself. In addition, the occipital condyle tends to be in a more dorsal position relative to the skull as a whole. This does not appear to be due to restructuring of the occiput so much as to the elongation of the quadrate ventrally. That is, the skull becomes deeper while the occiput remains more or less unaffected. Finally, the fused exoccipital-opisthotic element forms a broad bar joining the occiput to the quadrates on either side. Because of the increasing fenestration of the back of the skull, the entire occipital complex may appear to form a sort of Maltese cross, as in the rough sketch of the occiput of Chasmosaurus in Figure
6 (after Dodson (1996)). Here, much of the edge of the occiput is not in contact with any other bone. The lateral edges articulate with the quadrates, the very top is attached to the parietals, and the base rests more broadly on the basisphenoid.
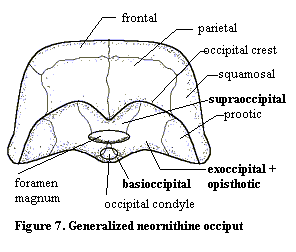 In birds (Figure 7), all the evolutionary labor of the archosaurs is undone. Broadly speaking, The bird skull is formed from the dinosaur skull by pushing all of the dorsal bones posteriorly. Conceptually, one might create bird from dinosaur by taking the heel of one's hand and kneading a theropod skull from front to back. The parietal is pushed onto the posterior face of the skull and may actually overhang portions of the occiput as an occipital crest. The foramen magnum and occipital condyle are pushed almost to the bottom -- even onto the ventral surface -- of the skull.
In birds (Figure 7), all the evolutionary labor of the archosaurs is undone. Broadly speaking, The bird skull is formed from the dinosaur skull by pushing all of the dorsal bones posteriorly. Conceptually, one might create bird from dinosaur by taking the heel of one's hand and kneading a theropod skull from front to back. The parietal is pushed onto the posterior face of the skull and may actually overhang portions of the occiput as an occipital crest. The foramen magnum and occipital condyle are pushed almost to the bottom -- even onto the ventral surface -- of the skull.
As in derived lepidosaurs, all of the elements in Figure 7 are fused in the adult skull. Accordingly, the divisions between individual ossifications are somewhat arbitrary. Note that the exoccipitals still participate in the occipital condyle. Since the bird condyle is rather small, and since one would expect an evolutionary premium on a smooth, even condylar head, it is hard to understand why evolution has preserved this 3-bone design.
As mentioned above, the synapsids develop a doubled occipital condyle, like neoselachians [2]. This development occurs at the level of the therapsids. Thrinaxodon, for example, clearly has two occipital condyles, although they remain -- perhaps somewhat awkwardly -- below the level of the foramen magnum. In eucynodonts, the condyles "migrate" dorsally so that they come to flank the foramen magnum. This arrangement completely prohibits lateral rotation of the head and thus requires a fully developed mammal-like atlas-axis complex (which permits a wide range of rotation between the first and second cervical vertebrae). The offsetting advantage is that shearing strains on the spinal nerves are minimized since the axis of flexion now passes through the spinal cord, rather than below it.
Additional Note
Two days after this essay was originally completed, I came across David Berman's Origin and early evolution of the amniote occiput (see Berman 2000)) -- which may be a greater comment on the shallowness of my original research than on the perversity of chance. In any case, this is a very fine paper combining a lot of careful, old-fashioned anatomy with a careful cladistic treatment of the results to yield an interesting and (despite the author's disclaimer) novel result. Crucial to this work is Berman's observation that the supraoccipital in tetrapods evolved independently at least twice, apparently from different embryonic structures.
ATW 010212
[1] Actually, in most typical early amniotes, the temporal opening would be less visible from this perspective, and the quadrate might be a smaller element. For the curious, this is a heavily edited, retouched, distorted and relabeled version of Proganochelys, an early turtle. The original is in Gaffney, ES & LJ Meeker (1983), Skull morphology of the oldest turtles: a preliminary description of Proganochelys quenstedi. J. Vert. Paleontol. 3: 25-28, vide Carroll (1988).
[2] Also like placoderms. However, the placoderm occiput is so unique that it is not really homologous to any of the structures addressed here. In placoderms, my (possibly deranged) understanding is that the craniospinal articulation has no condyle. The hinge between head and body is posterior to the braincase and consists of a usually doubled condyle joining the posterior cephalic and anterior thoracic plates.
checked ATW031117
{kind=link}