Sarcopterygii: Elpistostegalia
Metaxygnathus & Ventastega
Taxa on This Page
- Metaxygnathus X
- Ventastega X
Vikings, Ventastega and the Famennian Jaw (with a Long Discursion on Nordic History, Naval Architecture and Sundry Other Matters)
 With this page, we pass out of the Frasnian and into the somewhat different world of the Famennian elpistostegalians, including the first tetrapods. Like most other sorts of vertebrates, Elpistostegalia were hit fairly hard by the faunal turnover events at the end of the Frasnian. In fact, they seem to have suffered more than most, since no elpistostegalians are currently known from the earlier Famennian. The Famennian is a rather long period; and so this is a significant gap -- on the order of 10My. However, wherever they had been hiding, they emerged in droves in the Late Famennian and, this time, quickly attained a worldwide distribution. The identification of three new Famennian tetrapods in just the last three years, from China, France, and the United States, has confirmed that these Late Famennian forms were both diverse and widespread. Zhu et al. 2002) (Sinostega); Clement et al. (2004); Shubin et al. 2004).
With this page, we pass out of the Frasnian and into the somewhat different world of the Famennian elpistostegalians, including the first tetrapods. Like most other sorts of vertebrates, Elpistostegalia were hit fairly hard by the faunal turnover events at the end of the Frasnian. In fact, they seem to have suffered more than most, since no elpistostegalians are currently known from the earlier Famennian. The Famennian is a rather long period; and so this is a significant gap -- on the order of 10My. However, wherever they had been hiding, they emerged in droves in the Late Famennian and, this time, quickly attained a worldwide distribution. The identification of three new Famennian tetrapods in just the last three years, from China, France, and the United States, has confirmed that these Late Famennian forms were both diverse and widespread. Zhu et al. 2002) (Sinostega); Clement et al. (2004); Shubin et al. 2004).
Pending as always) new discoveries or a reclassification of old ones, Metaxygnathus and Ventastega are our last stop before the Tetrapoda. Since several of these near-tetrapods, as well as some of the early tetrapods, are jaw taxa, it is well to pay some attention to the details of the jaw. Fortunately, this requires little actual effort on our part, since we can pillage most most of the work done by Ahlberg & Clack (1998) in their excellent review of the topic. But, speaking of pillage, we will do so in our own fashion, by drawing an extended -- albeit somewhat strained -- analogy to the evolution of the Viking longboat.
To accompany us on this raid, we have invoked the image of a hypothetical Viking Ahlberg ancestor. This is patently unfair, since Prof. Ahlberg is Swedish, and has probably never even sacked a lab assistant, much less an English village. However, such trifling discrepencies should not be permitted to get in the way of the appropriate symbolism needed to invoke higher historical truths.
The Evolution of the Longboat
 What are those truths? Back in the mists of prehistory, the folks who would one day become the scourge of the North Sea and the North Atlantic (as well as France, Sicily and selected destinations in Byzantium), had all they could do to scourge each other. The typical raid of those times consisted of three or four young men quickly poling a hollow log across three kilometers of coastal marsh to bash their neighbors over the head with rocks, returning victorious (if at all) with the remains of some overaged goats, an underaged female, and perhaps a few portable metal nick-nacks. This highly informal system of dowery gradually evolved into a more collaberative culture when, sometime around 650 AD, the proto-Vikings noticed that other Northern European peoples frequently had younger goats -- and raised underaged females who were a shade less likely to murder their former captors and escape with all of the accumulated trinkets.
What are those truths? Back in the mists of prehistory, the folks who would one day become the scourge of the North Sea and the North Atlantic (as well as France, Sicily and selected destinations in Byzantium), had all they could do to scourge each other. The typical raid of those times consisted of three or four young men quickly poling a hollow log across three kilometers of coastal marsh to bash their neighbors over the head with rocks, returning victorious (if at all) with the remains of some overaged goats, an underaged female, and perhaps a few portable metal nick-nacks. This highly informal system of dowery gradually evolved into a more collaberative culture when, sometime around 650 AD, the proto-Vikings noticed that other Northern European peoples frequently had younger goats -- and raised underaged females who were a shade less likely to murder their former captors and escape with all of the accumulated trinkets.
However, long-distance "commerce" of this kind required better and more dependable transport. Now, unlike the master shipbuilders of the Mediterranean civilizations, the Viking shipwrights didn't think in terms of cargo tonnage, military logistics, or naval tactics. They thought in terms of hollow logs used to quickly deliver a few guys to a place where they could bash other people over the head, underaged females, etc. As things turned out, this led them to create hull designs markedly superior to those of their arguably more sophisticated neighbors.
The first Viking innovation was to add thin boards to the sides of the log and to carve a high-beaked bow. These boards were held on by struts running up the sides. Quite possibly, they were designed simply to keep everyone from getting soaked on the open sea. However, one could obviously get more space with less log and more cheap, light boards, if the boards were bent outward to extend the deck space. Over a few centuries of experimentation, the Vikings found that if they used a really, strong, flawless T-section keel in the center, with a really high, sharp bow for a cutwater, the rest of the vessel could be made almost entirely out of thin, flexible boards held together with lightweight struts braced on the keel. These lightweight materials had more "give" and less tendency to crack open in high seas, and the weight savings gave the longboats unmatched  speed and maneuverbility. Because of the hull configuration and the struts, any lateral forces on the hull were spread out across the surface and directed downward and inward "ventromedially") toward the strong keel. By the time of the Hypothetical Ahlberg -- at the height of Viking technological dominance in the Tenth Century -- the hollow log had been transformed into the long, low, alien-looking hull in the image. These were the fully-developed drakars or longboats, and their cargo-varrying cousins, the knarrs. They incorporated a shallow, sharp keelboard running the length of the ship, which gave much greater lateral stability at a minimal increase in draft. This design was so efficient and so deadly that it almost single-handedly forced Northern Europe to adopt the walled city, the castle, and the feudal system just to defend against the Hypothetical Ahlberg and his longboats.
speed and maneuverbility. Because of the hull configuration and the struts, any lateral forces on the hull were spread out across the surface and directed downward and inward "ventromedially") toward the strong keel. By the time of the Hypothetical Ahlberg -- at the height of Viking technological dominance in the Tenth Century -- the hollow log had been transformed into the long, low, alien-looking hull in the image. These were the fully-developed drakars or longboats, and their cargo-varrying cousins, the knarrs. They incorporated a shallow, sharp keelboard running the length of the ship, which gave much greater lateral stability at a minimal increase in draft. This design was so efficient and so deadly that it almost single-handedly forced Northern Europe to adopt the walled city, the castle, and the feudal system just to defend against the Hypothetical Ahlberg and his longboats.
But Wasn't This Supposed to be About Tetrapods?
Ah.... yes. Just so. In fact we were just getting to that point. Really.
Consider the skull of our constant companion in matters tetrapodomorph, Eusthenopteron. The head is a somewhat flattened, gently tapered cylinder, rigidly attached to the body. In short, its basic structure has quite a bit in common with the hollow log of the Viking ancestors. The main difference is that the fish swims under water, so that hydrodynamic considerations apply to the topsides as well as the hull. However, so long as the body form is more or less cylindrical, and the environment is symmetrical as well, these considerations are best served by a simple bullet or teardrop shape.
Body Form and Hydrodynamics
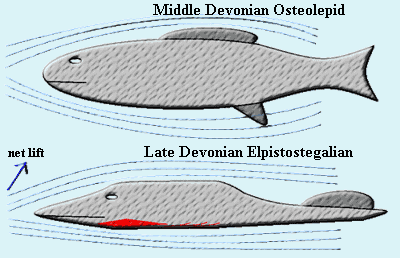 When we reach the Late Devonian Elpistostegalia, these assumptions are no longer true. First, these fishes have evolved significant dorsoventral asymmetry. The skull is now quite flat, and much flatter ventrally than dorsally. This fundamental assymmetry is probably applicable to the entire body form. The interclavicle continues the flat profile of the lower jaw onto the body. There are no ventral fins. Gastralia are commonly present which enforce this flat profile further down the body wall. Second, Late Devonian elpistostegalians are generally thought to have been air breathers, tolerating, if not requiring, frequent access to the surface.
When we reach the Late Devonian Elpistostegalia, these assumptions are no longer true. First, these fishes have evolved significant dorsoventral asymmetry. The skull is now quite flat, and much flatter ventrally than dorsally. This fundamental assymmetry is probably applicable to the entire body form. The interclavicle continues the flat profile of the lower jaw onto the body. There are no ventral fins. Gastralia are commonly present which enforce this flat profile further down the body wall. Second, Late Devonian elpistostegalians are generally thought to have been air breathers, tolerating, if not requiring, frequent access to the surface.
Notice what the change in body shape does to the aerodynamics or hydrodynamics, since they are still in the water). In underwater motion, the body asymmetry probably creates net lift, since the water must must travel further over the upper surface than the lower
-- just as in an the airfoil shape of an airplane wing. That's inconvenient for most fishes, but useful for an air breather. Further, the flatter ventral surface reduces frictional contact with the water when the fish is active on the surface. This profile also greatly improves maneuverability and shallow-water access for a surface swimmer.
This all sounds reasonable, but where's the evidence that the change in shape was actually used in this manner? The evidence is in its explanatory power. A fish of this sort will necessarilly encounter two sorts of hydrodynamic problems. First, how does it control the lift, particularly since the fins -- the usual control surfaces -- don't seem adequate for the task. Second, if our fish spends significant time on the surface, how can it maintain lateral stability without a keel or ventral fins?
 As to the lift problem, the skull of elpistostegalians is large, as well as flat. It is the curve of the head that creates the possibility of lift. What if we were to decouple the head from the body and allow it a degree of independent movement? Then the skull itself would become the control surface needed to control lift. This, of course, is exactly what occurred. The relaxation of intimate connections between the pectoral fins and the skull allowed the skull to move more freely and control the angle of the surfaces meeting water.
As to the lift problem, the skull of elpistostegalians is large, as well as flat. It is the curve of the head that creates the possibility of lift. What if we were to decouple the head from the body and allow it a degree of independent movement? Then the skull itself would become the control surface needed to control lift. This, of course, is exactly what occurred. The relaxation of intimate connections between the pectoral fins and the skull allowed the skull to move more freely and control the angle of the surfaces meeting water.
As to the second issue, modern shallow-draft vessels without keels often have removable centerboards -- short, roughly rectangular, boards which are sometimes paired (and not, despite the name, centered) if the vessel is broad in the beam. Centerboards don't have to do much, just move up and down as needed. Another, very similar, solution is to attach pontoons, as in an outriggger canoe. These pontoons can potentially also be adjusted and used as control surfaces for turning, although that isn't usual. The typical range of motion is restricted -- up, down, and folded out of the way when not needed. Again this is just the sort of structure we see developing in the earliest tetrapod limbs. The limbs are generally small, squarish or cylindrical, with a limited range of motion: up, down, and folded out of the way. In fact, there are further critical resemblances which we may touch on in connection with the digits of Acanthostega.
So, Are We Going to Talk About the Jaw or Not?
If you'd stop interrupting, yes we will.
If You'd Just Get to the Point, I Wouldn't Have To
 Very well. No need to shout. In spite of your insufferable abuse, we will proceed to the jaw.
Very well. No need to shout. In spite of your insufferable abuse, we will proceed to the jaw.
If the skull is the most significant control surface, and if these critters were often active on the surface, then the lower jaw becomes the most critical part of the system. It is here that the Viking longboat becomes a useful model. To be more exact, we must imagine a sort of double-keeled longboat, since the jaw is made up of two halves.
A Viking Catamaran?
Don't be absurd. Note quite. The two jaw rami are united anteriorly, but separate posteriorly. It's a single hull, but the keel bifurcates toward the stern. The principle is the same. But enough conceptual generalities. It is the structural details which matter.
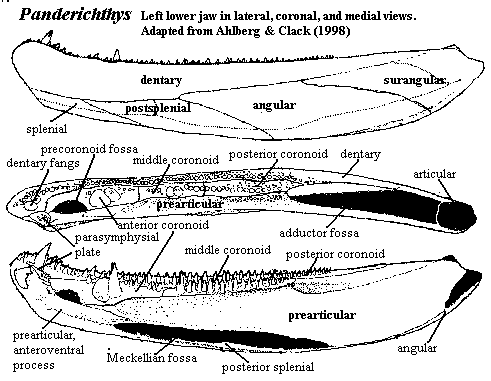
Ahlberg & Clack describe the jaw of Panderichthys as having: "a particular jaw morphology, where the coronoids are broad, and the anterior end of the jaw is deep and rather flat . In constructional terms, the anterior end of the jaw can be described as comprising an external plate of bone (formed from the dentary and splenial) with framework of solid strut-like elements (anterior coronoid,
'Jarvik's prearticular', Meckelian bone) applied to the inner face." [AC98: 40].
This is, to a first approximation, the elaborated hollow log of the proto-Vikings. The anterior jaw is a blunt, boxy affair, made up of thick, relatively massive bones. This design, while relatively clumsy, does have the advantage of allowing the jaw to bear large fangs in all kinds of places. However fangs may have ben used in feeding, they certainly represent points of considerable potential stress; but, as on a barge or a scow, the placement of stress points isn't particularly critical.
 The anterior jaw in the Frasnian models, Elginerpeton and Obruchevichthys, has a comparatively narrow surface meeting the water, mostly by virtue of being tapered and very long. The basic construction is still relatively flat and boxy, as described by Ahlberg & Clack. However, we also see the beginnings of more fundamental changes. The parasymphysial plate becomes longer and more substantial. From the limited available data, it seems likely that the lateral fangs were being de-emphasized, while the parasymphysial plates were taking on a greater role in bearing both fangs and teeth. The external profile becomes rounder and smoother, like the evolving Viking longboat, and there appear to be some modest development of the splenial in the nature of a keel. See, e.g. the ventral jaw surfaces in Livoniana and Obruchevichthys.
The anterior jaw in the Frasnian models, Elginerpeton and Obruchevichthys, has a comparatively narrow surface meeting the water, mostly by virtue of being tapered and very long. The basic construction is still relatively flat and boxy, as described by Ahlberg & Clack. However, we also see the beginnings of more fundamental changes. The parasymphysial plate becomes longer and more substantial. From the limited available data, it seems likely that the lateral fangs were being de-emphasized, while the parasymphysial plates were taking on a greater role in bearing both fangs and teeth. The external profile becomes rounder and smoother, like the evolving Viking longboat, and there appear to be some modest development of the splenial in the nature of a keel. See, e.g. the ventral jaw surfaces in Livoniana and Obruchevichthys.
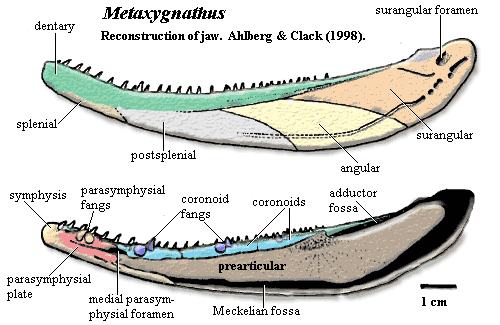
The Famennian versions go a considerable step further. The engineering constraint which confined the earlier models to a boxy design was the necessity of accommodating the gular and branchiostegal rays below the jaw. In the Famennian forms, these bones had been eliminated, or at least restricted to the more posterior sections of the jaw. The entire curve of the jaw now takes on a boat-like form, and the anterior forms a proper, solid prow formed by the large parasymphysial plate and medial lamina of the splenial -- a general strengthening and centralization of the anterior jaw. Daeschler 2000). Already in Metaxygnathus, we can see the splenial keel extending posteriorly, closing the Meckelian fossa. This process of "zippering up" the fossa from anterior to posterior is one which continues through the early tetrapods. In the tetrapods, the dentary is recruited to assist, and eventually to replace, the splenial. Eventually, this sharp, unitary keel replaces the Meckelian bone as the structural foundation of the jaw, and the Meckelian is no longer ossified along the main part of the jaw. As this "tetrapod keel" is established, the medial and lateral dermal bones are gradually freed up to become lighter and more flexible, to develop specialized forms, or simply to disappear.
So What's The Bottom Line?
 The keel is, so to speak, the bottom line -- by definition. It is also a suitable centerpiece for our demonstration here. What we conclude is that the main outlines of jaw evolution in the Late Devonian can be explained by reference to locomotion alone, and in accordance with historically verifiable principles of naval architecture. Obviously, no matter how sleek and maneuverable the jaws may be, they are of no use if they cannot feed the organism which bears them. Therefore, we assume that considerations of prey capture and consumption were also important.
The keel is, so to speak, the bottom line -- by definition. It is also a suitable centerpiece for our demonstration here. What we conclude is that the main outlines of jaw evolution in the Late Devonian can be explained by reference to locomotion alone, and in accordance with historically verifiable principles of naval architecture. Obviously, no matter how sleek and maneuverable the jaws may be, they are of no use if they cannot feed the organism which bears them. Therefore, we assume that considerations of prey capture and consumption were also important.
Nevertheless, we are tempted to assert that the most significant selective forces at work on Late Devonian stem tetrapods were similar to those facing the Hypothetical Ahlberg. His technological development was conditioned by the availability of the necessities of life -- goats, girls, and goodies, in no particular order -- which a numerically and economically superior culture had conveniently deposited at diverse locations along the coastline. The strategy adopted by the Hypothetical Ahlberg was to use his superior mobility and initiative to acheive locally overwhelming force.
Our own hypothetical ancestors in the Devonian seem to have evolved a comparable technology. Did they do so for comparable reasons? That is, did they live in an environment in which littoral prey was plentiful and easily consumed, if the predator were quick and didn't overstay its welcome? The nature of that environment is what we will take up eventually) in connection with the first tetrapods.
ATW050516. Text public domain. No rights reserved.
Metaxygnathus: M. denticulus Campbell & Bell, 1977. Isolated jaw ramus.
Range: Famennian of Australia (NSW)
Characters: jaw slender & strongly curved, generally similar to Acanthostega [AC98]; posterior dentary narrow & splint-like [AC98]; ventral margin of dentary beveled to a thin edge & loosely attached to angular & surangular [AC98]; surangular with large ventrally positioned foramen [AC98$]; medial parasymphysial foramen large [AC98]; lateral parasymphysial foramen unknown [AC98]; preparticular reaches medial parasymphysial foramen [AC98]; prearticular sutures anteroventrally with medial lamina of splenial which wraps around prearticular [AC98]; prearticular with poorly developed dorsal denticulated strip [AC98]; probably a wide Meckelian fossa with ossified Meckelian [AC98]; prearticular approaches but does not contact angular [AC98]; parasymphysial plates with fang pairs & tooth row [AC98]; 3 coronoids [AC98]; dentary with anterior fang [AC98]; no accessory dentary tooth row [AC98]; coronoids with fangs inside tooth row [AC98]; bones very lightly ornamented [AC98];
References: Ahlberg & Clack (1998) [AC98]. ATW050502.
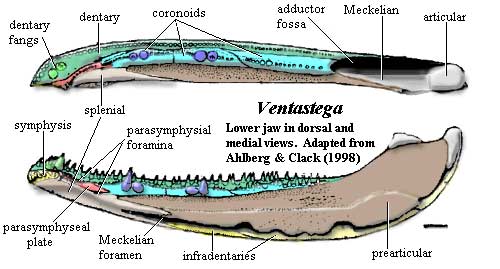
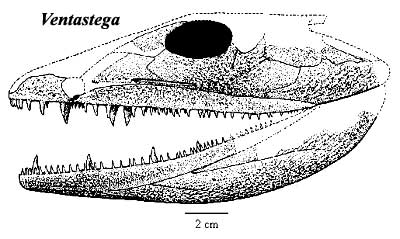 Ventastega: V. curonica Ahlberg et al., 1994.
Ventastega: V. curonica Ahlberg et al., 1994.
Range: Famennian of Latvia
Characters: paired median rostrals [AL04]; nasals not sutured together, with fontanelle (gap) in midline [C03] [AL04]; dentary "chamfered" (ventral margin of dentary beveled to a thin edge & loosely attached to angular & surangular) [AC98] [D00] [R+03] [AL04]; dentary dorsoventrally thin posteriorly [D00]; dorsal edge of splenial contacts anterior coronoid [D00]; Meckelian ossified throughout [AL04]; articular placed well above tooth row [AC98]; shallow, widened coronoid region with roughly square-shaped cross-section [D00]; lateral and medial parasymphysial foramina present [AC98]; parasymphysial plate with tooth row [R+03]; ventral margins of prearticular & infradentaries held together by strip of Meckelian bone [AC98]; prearticular contacts angular, covering Meckelian [AC98]; vomer & palatine lack accessory row of denticles [C02a]; palatal fangs present [C02]; fang pairs and tooth row on anterior and middle coronoids [C02a] [R+03]; coronoid fangs in tooth row [A95]; tooth row on posterior coronoid [R+03]; denticles on prearticular confined to narrow strip [AC98]; clavicle with broad triangular base and narrow ascending lamina [C02]; interclavical large & rhomboidal [C02] [AL04]; ribs short & slender [AL04]; ilium slender, with posterodorsally directed postiliac process & no iliac canal [AL04]; tail fin with fin rays [AL04]; fresh water environment? [LG04].
Image: Ventastega from the Natural History Museum site, © The Royal Society of London.
Notes: [1] Clack [C02a] notes that Ventastega is very similar to Acanthostega in cranial anatomy, but quite different post-cranially.
References: Ahlberg 1995) [A95]; Ahlberg & Clack (1998) [AC98]; Ahlberg & Luksevics (2004) [AL04]; Clack 2002) [C02]; Clack 2002a) [C02a][1]; Clack (2003) [C03]; Daeschler (2000) [D00]; Long & Gordon (2004) [LG04]; Ruta et al. (2003) [R+03].
