
Archegosaurus. Early Permian period, north-central Pangea Europe). Length 1.5 meters
| Temnospondyli | ||
| The Vertebrates | Temnospondyli |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Tetrapoda
├─┬─Lepospondyli
│ └─Reptiliomorpha
│
└─Temnospondyli
└─┬─Edopoidea
│ ├─Edops
│ └─Cochleosauridae
└─┬─Dendrerpeton
├─Dvinosauria
└─┬─Euskelia
│ ╞═Dissorophoidea
│ │ └─Lissamphibia
│ └─Eryopoidea
└─Stereospondyli
├─Rhinesuchidae
└─┬─Lydekkerinidae
├─┬─Plagiosauroidea
│ └─┬─Rhytidosteidae
│ └─Brachyopoidea
└─┬─Capitosauria
└─Trematosauria
├─Trematosauroidea
└─Metoposauroidea
|
Contents
Index |
|
|
Archegosaurus. Early Permian period, north-central Pangea Europe). Length 1.5 meters |
The Temnospondyls were an extremely diverse and succesful group of "labyrinthodont" tetrapods. They represent one of the three primary lines of labyrinthodont evolution, the other two being the reptilomorphs and lepospondyls. Although some temnospondyls were completely aquatic and even had external gills as adults, others probably became almost as terrestrial as reptiles, returning to water only to lay their eggs. The Temnospondyli evolved in the Early Carboniferous. Most Paleozoic forms became extinct before or during the Lopingian (Late Permian). However two aquatic Gondwanan clades, the dvinosaurs and stereospondyls, survived and even prospered in the early Mesozoic. The youngest known temnospondyl is Koolasuchus from the Middle Cretaceous (Barremian) of Australia.
Temnospondyls had, for the most part, relatively large heads, with rather flat, akinetic skulls. The shape of the skull was quite variable, and often distinctive. Some, like Archegosaurus, had the mouth drawn out into the classic "longirostrine" shape associated with reptiles which hunt small fish. Others, like Gerrothorax, had bizarre, wide, parabolic heads like galeaspids or other "cornuate" jawless fish. Some, Laidleria for example, had very flat, triangular heads with no easy parallels in any other vertebrate group, before or since.
In contrast, the temnospondyl post-cranial body plan was conservative and fairly unspecialized. The body proportions varied considerably from one group to another. As a result, we may suspect that some tended toward an eel-like style of swimming or an undulating, almost legless type of locomotion on land. Others had relatively stiff, massive bodies and presumably used their limbs for both land and water locomotion. However, no temnospondyl group developed any limb specializations as in the reptilomorph lineage. The temnospondyl fore-foot had only four toes, the hind-foot five. The vertebral structure was essentially of the primitive, intercentra and pleurocentra sort, as in the early Devonian tetrapods and osteolepiform lobe-finned fish. However there was also some variation in vertebral structure, a distinction being made between the ancestral, ratchitomous condition that characterized the majority of temnospondyls, and the more specialized sterospondylous condition of the later, aquatic, Triassic types.
Colosteid |
Dendrerpeton |
Branchiosaur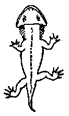 |
Eryops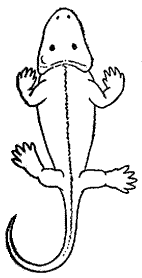 |
Mastodonsaurus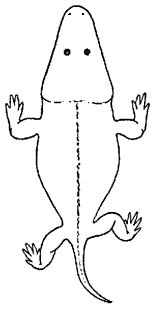 |
| Early to late Carboniferous | Late Carboniferous-early Permian | Triassic | ||
Perhaps the single most successful group of non-amniote tetrapods, the temnospondyls have a long and diverse history.
Sometime in the Early Carboniferous, a lineage of early tetrapods (primitive labyrinthodont amphibians in pre-cladistic speak) gave rise to a new and important clade (or grade), the temnospondyls. Like any successful group of animals, they radiated quickly into many new forms, some of which became progressively more aquatic, while others spent more and more time on land, and others that continued a typically amphibious lifestyle.
They included not only a large element of terrestrial and fresh-water aquatic faunas of the Carboniferous and Early Permian, and semi-aquatic and aquatic from the Middle Permian to the Late Triassic, and eventually (beginning in the Triassic), modern amphibians.
The first proto-temnospondyl - like species were fierce scaly aquatic animals, rather like a cross between a salamander and a moray eel. These were the Colosteids. From these arose medium to large-sized swamp dwellers known as Baphetids, and from these in turn, perhaps via forms like Eucritta, the first true temnospondyls.
The early temnospondyls therefore arose out of the original tetrapod radiation of the Tournaisian and Visean (early Carboniferous) epoches.
They included a number of similar, broadly crocodile-like forms, about a meter to one and a half meters in length, but quickly diversified intoa large range of forms. These animals were at their height in the tropical coal-swamp biome of the middle Carboniferous (late Visean epoch), where they shared the waterways with even bigger and fiercer eel-like animals. Already co-existing with the Colosteids during the late Visean were the massive Eryops and the small and rather inoffensive-looking Dendrerpeton. Unlike the Colosteids, these were terrestrial, or at least semi-aquatic, with larger legs stiffer bodies.
 Amphibamus grandiceps, a dissorophoid - Late Carboniferous (Moscovian) of Illinois. This animals is considered to have been close to the ancestry of moidern amphibains. Length about 20 cm. Life reconstruction, by Arthur Weasley (Wikipedia) |
The heyday of the Paleozoic temnospondyls was during the late Carboniferous and early Permian periods. During this time there was a rich diversity of these creatures, some with huge heads and armour plates, and adapted to a terrestrial existence, such as the Dissorophid Cacops, others with strong robust limbs and large heads, but still semi-aquatic and rather like a cross between a crocodilian and an enormous frog (Trematops, Eryops), others aquatic with long bodies and armoured with a protective coating of scales (Trimerorhachis) while others again retained external gills even as adults and were unable to leave the water (Branchiosaurus).
The increasing aridity of the late Permian world, brought about the single landmass (the Pangean supercontinent) and disruption of rainfall patterns, and the sucess of the reptiles, was not the best news for these large tetrapods, and they retreated to the swamps, ponds and river courses. Originally these Late Permian temnospondyls had a crocodile-like apperance and were probably still quite capable of climbing out on land (Actinodon, Archegosaurus, Platyoposaurus), but these transitional types were soon replaced by their descendents, the large to very large, completely aquatic stereospondyls. One lineage of stereospondyls, the Trematosauridae, actually took up a life in the oceans, the only stem tetrapods ever to have done so. The Trematosaurs underwent quite an evolutionary radiation, and the various genera are distinguished by the proportions and shape of their skulls.
During this same period, other Stereospondyl lines continued to thrive, but also evolved in more extreme directions. Because these creatures lived their entire lives in water (although they were probably able to climb out on land for short periods) their skeletal structure became increasingly cartiliginous, and their heads increasingly large and flat. Among the Capitosaurids and Mastodonsaurids the head became so large and flat in fact that it is difficult to see how the animals could have moved their lower jaws. Presumably, they only opened their mouths by titling back the whole head.
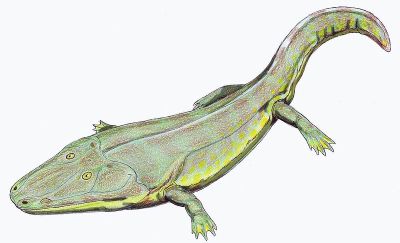 Cyclotosaurus, a capitosaur, Triassic (Carnian) of Europe. Capitosaurs were one of a several groups of large flat-headed forms that inhabited Triassic watercourses. Length 2 to 4 meters. Life reconstruction, by Dmitry Bogdanov (Wikipedia) |
Both large and small aquatic Temnospondyls continued as an important part of the freshwater ecosystem right up until the end of the Triassic period, even co-existing with the formidable crocodile-like phytosaurid reptiles. Important at this time were the Metoposaurs of Laurasia (Europe and America), which evolved from completely different ancestors to look strikingly like the Capitosaurs. Indeed, the only easy distinction is that the Metoposaurs had eyes further forward on the skull.
The terminal Triassic extinction killed off all the big temnospondyls, along with the phytosaurs. Only a few, short-headed stragglers making it through to the Jurassic the Brachyopoids. These survived in China and Australia (the easternmost parts of north and south Pangea respectively), perhaps protected by geographical isolation. Safe from enemies, they grew to be as big as Capitosaurs. The last temnospondyls were two and a half to three meter long giants who sought refuge in the Cretaceous polar rift valleys of south-east Gondwana, where the climate was too cold to support crocodiles, their main predators.
MAK090710
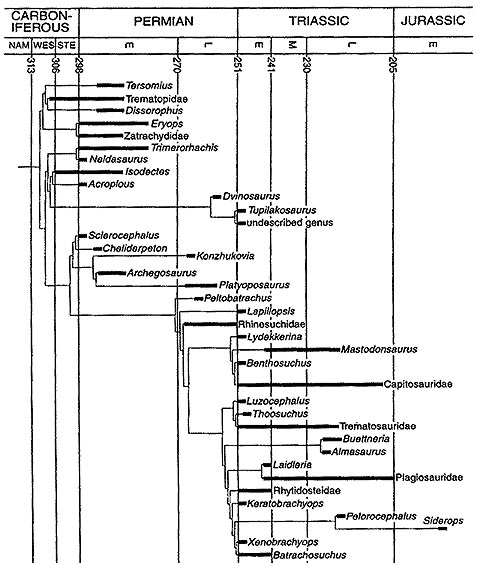 Traditionally, there were only two temnospondyl suborders, the Ratchitomi and the Stereospondyli, which were determined according to vertebral type, and these are mentioned in the older books. Romer provided a third suborder, the fish-eating marine-going trematosaurs. Other paleo-tetrapod authorities such as Nilsson and Panchen removed the short-headed Peltobatrachidae and Plagiosauridae from the Brachyopoids and placed them in a seperate order. Later writers like Carroll and Benton did away with the ratchitome - sterospondyl classification altogether, arguing that the sterospondyls were an artificial (polyphyletic) group, the stereospondylous condition having evolved seperately a number of times.
Traditionally, there were only two temnospondyl suborders, the Ratchitomi and the Stereospondyli, which were determined according to vertebral type, and these are mentioned in the older books. Romer provided a third suborder, the fish-eating marine-going trematosaurs. Other paleo-tetrapod authorities such as Nilsson and Panchen removed the short-headed Peltobatrachidae and Plagiosauridae from the Brachyopoids and placed them in a seperate order. Later writers like Carroll and Benton did away with the ratchitome - sterospondyl classification altogether, arguing that the sterospondyls were an artificial (polyphyletic) group, the stereospondylous condition having evolved seperately a number of times.
However, the relationships between the different temnospondyl families, and sometimes even the question of which family goes in which superfamily, remained very controversial. Many of these problems in temnospondyl taxonomy were addressed by Yates & Warren (2000) in comprehensive cladistic analysis from which the following cladogram is taken.
In this diagram, the geological period is shown on the top. The known occurance of taxa (genera and families) in the fossil record is indicated by solid bars. Thin lines indicate hypothetical ancestry (cladistic techniques forbid deriving one known monophyletic genus or family from another, rather, both are derived from a hypothetical "most recent common ancestor", which gives the branching pattern as shown above).
Several things need to be said about this diagram. First, not every temnospondyl taxon is included. Many known types are not shown here. However, most of the important forms are shown. Secondly Yates and Warren revive the monophyletic Trematosauria and Stereospondyli, but replaced the Ratchitomi with the clades Euskelia and Limnarchia. And third, not everyone agrees with these particular phylogeny. A common alternative for example is to have the Dvinosauria as a very early and quite distinct clade, prior to the Euskelia and the higher "Limnarchia" (several of the dendrograms displayed here show this option)
Temnospondyli: Dendrerpeton.
Range: from the Early Carboniferous
Tetrapoda*: Lepspondyli + Reptilomorpha) + *: Eucritta + (Edopoidea + (Dendrerpeton + (Euskelia + Limnarchia))).
20- 300 cm; large heads with akinetic skulls; skull triangular to parabolic, or longirostrine; skull heavily ornamented; sensory line grooves may be present; $ palatal tusks common; $ very large interpterygoid fenestrae; tendency to develop additional cranial bones along midline (e.g. internasal, interfrontal, interparietal); "otic notch" frequently present, but probably did not support tympanum; "vertebrae 'rhachitomous' (with a large, dorsal, crescentic intercentrum and a small, dorsal, paired pleurocentrum) to stereospondylous without an ossified pleurocentrum, although this element may be retained in a cartilaginous state)" [from Temnospondyli]; $ ornamented scapula; digits 4 (manus), 5(pes); amphibious or aquatic; some forms even with probable external gills.
Links: Dendrerpeton and Joggins, Nova Scotia; Temnospondyli; Basal Temnospondyli; Historický vývoj (Czech); Phylogeny and Apomorphies of Temnospondyls. 010806.
Discussion: 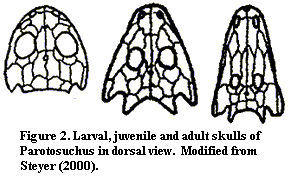 One of the nice things about temnospondyls is that there are a great many of them. Not only did they manage to diverge into a great many species, but the more successful groups are known from numerous specimens. Thus, it is possible to examine aspects of this taxon which cannot normally be studied in paleontological material -- such as developmental biology. In several cases, there are so many specimens known that it has become possible to piece together the ontogenesis of temnospondyls. Temnospondyls are normally thought of as having three life phases: (a) a larval stage, probably aquatic and gill-breathing, (b) a juvenile transition phase; and (c) an adult form. However, in comparing species, taxonomists have largely restricted themselves to the adult morphs.
One of the nice things about temnospondyls is that there are a great many of them. Not only did they manage to diverge into a great many species, but the more successful groups are known from numerous specimens. Thus, it is possible to examine aspects of this taxon which cannot normally be studied in paleontological material -- such as developmental biology. In several cases, there are so many specimens known that it has become possible to piece together the ontogenesis of temnospondyls. Temnospondyls are normally thought of as having three life phases: (a) a larval stage, probably aquatic and gill-breathing, (b) a juvenile transition phase; and (c) an adult form. However, in comparing species, taxonomists have largely restricted themselves to the adult morphs.
Recently, Jean-Sébastien Steyer, a graduate student at the Muséum National d'Histoire Naturelle in Paris, asked himself what would happen if one didn't discard all of the developmental data. Steyer 2000). Rather than (literally!) throwing out the babies with the bath water, Steyer set out to compare six different in-group taxa (plus Dendrerpeton as the out-group) whose developmental course was particularly well charted. The particular taxa and results were not particularly novel in this case, and were fully consistent with Yates & Warren 2000). As intended by Steyer, the point of real interest was not the results, but the method -- a method which has some very exciting possibilities.
In brief, Steyer scored the same 40 characters for each stage of each species studied, in effect treating a larval Apateon as separate species from the juvenile and adult forms for purposes of scoring. The scores were generally based on Dendrerpeton as the out-group, although there were some missing data and a few other exceptions. Separate cladograms ("ontotrees") were then generated for each morph across all species. Finally, a total evidence cladogram was generated, using all three sets of data. Interestingly, the three ontotrees were quite different. However, the combined data generated a topology which was identical with the larval cladogram and was quite robust (CI = 0.72).
Steyer was kind enough to discuss some of the fine points of the technique with me. In the process, I came to the conclusion that this method is at least as powerful as Steyer hopes it will be. In fact, one cannot easily think of a technique more likely to resolve the intractable deep nodes in the neornithine or mammalian radiations than phylogenetic taxonomy based on developmental biology. Haeckel's Law is not a "law," but it is certainly a frequent observed phenomenon. That is, we have good reason to hope that a systematic analysis of ontogeny will at least help us recapitulate (actually, recalculate) phylogeny. Conveniently, the living embryos of those species are around for all to see, so the work may be much more detailed -- and much easier -- than piecing together the shattered dermal bones of Carboniferous tetrapods.
But, first, there are some surprisingly thorny theoretical questions to be resolved. For example, just what is a synapomorphy in this context? Normally, the textbook answer is: a shared derived characteristic of a clade. Like a family recipe for lamb curry, or a tendency to practice obscurity for its own sake, some things mark a family of humans or other vertebrates and can duly be scored to produce cladograms. But what if say that our family is the one with beards? "So what?", you say, "Lots of people have beards." But no, I explain, in my family everyone has a beard: women, small children, parakeets, the lot. Thus the question of synapomorphy is more complex than presence or absence. It matters when those characters appear. And, if otherwise normal small children have beards in a family, would I score the beards of adult males in that family as a plesiomorphic feature of normal humans (state 0), or as the continuation of a peculiar apomorphy of my family?
Let us take a look at a very concrete example. In Figure 2, it is evident that the Parotosuchus has a large otic notch as a larval form, a notch which is still quite noticeable as a juvenile. This is a derived state. Dendrerpeton, the outgroup for almost all purposes, has little if any otic notch as a larva or juvenile. However, the adult morph of Parotosuchus has a small, narrow otic notch which looks very similar to the small, narrow otic notch of the adult Dendrerpeton. Steyer scores the otic notch of Parotosuchus 1 (larval), 1 (juvenile), and 0 (adult). But, if the otic notch of the adult is developed from the open notch of the juvenile in Parotosuchus, rather than from a notchless skull as in Dendrerpeton, is this really the same state or a developmental homoplasy?
Perhaps the latter is a better choice. That is, "once derived, always derived" is a reasonable rule of thumb. We are essentially dealing with developmental vectors, not the usual scalar quantities of a typical cladistic study. Each stage-specific character not only has a definite description, but also a developmental direction. The adult otic notch looks the same, but it developed from the opposite direction. Once derived, a character does not "underive." In normal cladistics, we may discover, ex post, that a what appeared to be a plesiomorphy was actually a reversal. But the calculation has already been performed. Here, we know ex ante that the notch is developmentally different from the notch of Dendrerpeton, so it should be scored differently from the beginning, and before the tree is calculated.
But Steyer may yet have the better argument. Consider the example of the beards. What if the beards of the women and children and, of course, the parakeets) are developmental neomorphs brought about by some unique hormonal aberration, while the beards of adult males are the result of the normal pattern of development reasserting itself? How can I justify scoring the adult males as derived just because the children are?
Ultimately, it may be a judgment call. Or one might separately score the direction and value, so that the 1 -> 0 transition itself is given a score different from the 0 -> 0 development of Dendrerpeton. The problem is that this practice clearly violates the rule of independence. That is, the scoring of transitions depends in a simple way on the underlying states, with the result that the states may effectively be double-counted. This may distort the results significantly. In Steyer's 3-stage study, for example, scoring transitions would essentially require us to count the middle, juvenile state twice as often as the larval and adult forms (i.e. la->ju and ju->ad). This is a form of weighting which has no obvious theoretical justification.
This is a powerful technique with a great deal of promise, but working out exactly how to use it is not an easy proposition.
References: Steyer (2000); Yates & Warren (2000).
checked ATW060327
{kind=link}
{kind=link}