 The Standard Condition of the jugal is shown at right. In this case, our model is not Bob, our basal amniote. Instead, we have selected a slightly more primitive relative, Eoherpeton. The image is relabeled from Smithson (1985), vide Clack (2002).
The Standard Condition of the jugal is shown at right. In this case, our model is not Bob, our basal amniote. Instead, we have selected a slightly more primitive relative, Eoherpeton. The image is relabeled from Smithson (1985), vide Clack (2002).| Bones: Dermal Bones | ||
| Vertebrates | Orbital Series: Jugal |
| s ("Cladograms") |
||||||
| Vertebrates Home | Vertebrate | Vertebrate | Bones | Time |
|
Orbital Series
|
Unlike many of the odder bones with which we have been required to deal, the jugal is almost an open book. It is generally easy to locate and identify. Its structural role is straightforward; and its position and properties have been quite stable over time. Each of these rules is subject to some exceptions; but, by and large, we can coast on this one.
The Standard Condition of the jugal is shown at right. In this case, our model is not Bob, our basal amniote. Instead, we have selected a slightly more primitive relative, Eoherpeton. The image is relabeled from Smithson (1985), vide Clack (2002).
At least at this stage, the jugal almost always underlies the orbit, and contacts the lacrimal anteriorly. The relationship of jugal and maxilla is variable, although they are almost always in contact. Generally, the jugal passes dorsal to the maxilla to terminate anteriorly on the lacrimal. Romer (1956). Sometimes, as in Eoherpeton, the maxilla and quadratojugal exclude the jugal from the lower margin of the skull. More often, the jugal forms the ventral margin and separates the quadratojugal and maxilla. The jugal's contacts dorsally are a bit more variable due to the variety of temporal fenestration seen in the amniotes. In general, its dorsal partners are the squamosal and either the postfrontal or postorbital.
Among amniotes, it is useful (but not always accurate) to think of the jugal as a three-pronged bone centered posteroventral to the orbit. From this perspective, the three rami are : (1) an anterior or suborbital process which passes under the orbit and terminates at the lacrimal and/or maxilla; (2) a posterior or subtemporal process which forms the bar (if any) under the temporal fenestra (if any) and terminates at the quadratojugal; and (3) a dorsal or postorbital ramus which forms part or all of the postorbital bar and terminates on the postfrontal, postorbital, or whatever. In this avatar as a trimurti, the jugal obviously plays an important role, defining and limiting the degree of the skull's lateral flexibility, as well as joining together the disparate portions of the dermal skull at this central point. This, in fact, may be the source of its name, as iugare is the Latin meaning to bind together, as in a marriage or a team of horses (iugales).
 In a sense, the jugal traces its ancestry to some of the oldest specialized dermal bones, for practically since vertebrates had eyes, they have had small, specialized, dermal bones around and behind the eye to protect it from damage or deformation. Thus, the arandaspid Sacbambaspis is believed to have had a sclerotic ring. Janvier (1996: 86), And the anaspid Rhyncholepis had a small circle of dermal bones surrounding the orbit. Janvier (1996: 101). Many placoderms had one or both, and it is from this circumorbital series that the jugal seems to have derived.
In a sense, the jugal traces its ancestry to some of the oldest specialized dermal bones, for practically since vertebrates had eyes, they have had small, specialized, dermal bones around and behind the eye to protect it from damage or deformation. Thus, the arandaspid Sacbambaspis is believed to have had a sclerotic ring. Janvier (1996: 86), And the anaspid Rhyncholepis had a small circle of dermal bones surrounding the orbit. Janvier (1996: 101). Many placoderms had one or both, and it is from this circumorbital series that the jugal seems to have derived.
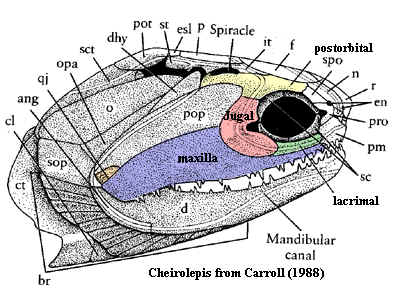
Already in the Acanthodii, one of the circumorbital bones, at the posteroventral corner of the orbit, has become notably enlarged and carries the infraorbital sensory line on its surface -- the mark of the jugal in fish. By time we reach Cheirolepis, our usual starting point for dermal bones, the jugal is relatively large and beginning to take on a structural role beyond the simple job of holding the eye in place. However, at this point, the jugal has no relation to the quadratojugal, nor any role in the ventral margin of the skull.
Actinopterygii: Subsequent to its development in Cheirolepis, the jugal has not had a distinguished career among the ray-finned fish. Because of the fundamental reorganization of the jaw in this group, the maxilla is short and mobile in all derived forms. Consequently, there is little need for a stout bone in this position. The jugal simply blends back in to a series of undistinguished and almost indistinguishable suborbitals that serially absorb the strains imposed by movement of the maxilla against the relatively stationary preopercular, posteriorly, and the skull table, dorsally.
Sarcopterygii: By contrast, the maxilla is relatively immobile in sarcopterygians. Indeed, in the Actinistia (e.g. the coelacanth) and Dipnoi (lungfishes), the upper jaw is completely immobile and ultimately fused to the braincase. In these forms, it is the maxilla which disappears and the jugal (or other suborbitals) which provide the dermal covering of the anterior part of the upper jaw. Along the main line of sarcopterygian evolution, the case was not so extreme. The jugal became an important arbiter between the conflicting claims of the jaw, the orbit and, most importantly, the squamosal. This last was a novel bone which was beginning to contend with the traditional opercular series for domination of the lateral skull.
In our own ancestors, the Rhipidistia, the maxilla is slight, but the jaw, maxilla and all, is considerably elongated -- presumably to a point at which it became impractical to rely on an immobile shoulder girdle for ultimate posterior support. But that is another story. Here, the point is that the long jaw and skull made support for the postorbital skull problematic, particularly given the little sliver of maxilla and the growth of more massive jaw adductors. As a result, the jugal becomes a massive plate stretching back through the enlarged former territories of the maxilla and preopercular to reach a similarly behemoth quadratojugal.
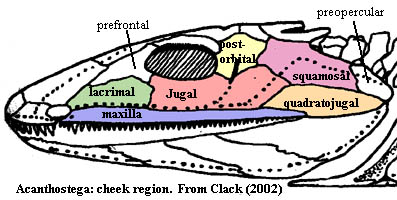 Tetrapoda: Thus, by the Late Devonian Acanthostega,the skull looked somewhat like the figure at right from, Clack (2002). Note that the squamosal has been interposed anterior to the rapidly shrinking preopercular. Note the shape of the jugal. If we consider its home range as a circumorbital bone, it almost appears as if the jugal had been doubled or repeated, with the two halves joined. This impression is only reinforced by comparing the respective courses of the sensory lines through the jugal in Cheirolepis and Acanthostega.
Tetrapoda: Thus, by the Late Devonian Acanthostega,the skull looked somewhat like the figure at right from, Clack (2002). Note that the squamosal has been interposed anterior to the rapidly shrinking preopercular. Note the shape of the jugal. If we consider its home range as a circumorbital bone, it almost appears as if the jugal had been doubled or repeated, with the two halves joined. This impression is only reinforced by comparing the respective courses of the sensory lines through the jugal in Cheirolepis and Acanthostega.
Temnospondyls: Perhaps there is something to the last thought, since the orbit seems to roll back and forth on the jugal within basal tetrapods and early temnospondyls. In Dendrerpeton, the orbit is well forward on the jugal, narrowing the jugal almost to a point, so that it makes only slight contact with the lacrimal. In Eryops, the orbit is more dorsal and posterior. Here, the jugal broadly contacts, not just the lacrimal, but also the prefrontal as in Acanthostega. The latter scheme, involving a jugal extending well anterior to the orbit, is asserted to be a synapomorphy of the Stereospondylomorpha. Yates & Warren (2000). However, some Triassic temnospondyls, such as Laidleria, exhibit the pattern of Dendrerpeton, with the jugal and prefrontal broadly sutured, and the orbits well back on the skull. Warren (1998). Perhaps it may be more accurate to suppose that the position of the orbit changes, while the form and relationships of the jugal remain rather constant. Interestingly, in few, if any, temnospondyls does the jugal contribute to the ventral margin of the skull. This appears to be a more stable phylogenetic character and is less dependant on the relative position of mutable markers like the position of the orbits.
 Lepospondyli & Lissamphibia: The pattern of the jugal in lepospondyls is generally conservative, more or less like Acanthostega except that the jugal finally reaches the ventral margin of the skull in some forms, such as the microsaurs Hapsidopareion and Rhynchonkos and the Nectridia. (Carroll 1988) A "normal" jugal is present also in the Early Jurassic gymnophionan Eocecilia. Jenkins & Walsh (1993). However in the Late Jurassic Karaurus, an early salamander, and in all later Lissamphibia, the jugal is usually absent altogether. While the details are obscure, to us at least, this development may relate to the development of specialized structures to project the tongue, with emargination of the ventral skull margin to accommodate the necessary musculature.
Lepospondyli & Lissamphibia: The pattern of the jugal in lepospondyls is generally conservative, more or less like Acanthostega except that the jugal finally reaches the ventral margin of the skull in some forms, such as the microsaurs Hapsidopareion and Rhynchonkos and the Nectridia. (Carroll 1988) A "normal" jugal is present also in the Early Jurassic gymnophionan Eocecilia. Jenkins & Walsh (1993). However in the Late Jurassic Karaurus, an early salamander, and in all later Lissamphibia, the jugal is usually absent altogether. While the details are obscure, to us at least, this development may relate to the development of specialized structures to project the tongue, with emargination of the ventral skull margin to accommodate the necessary musculature.
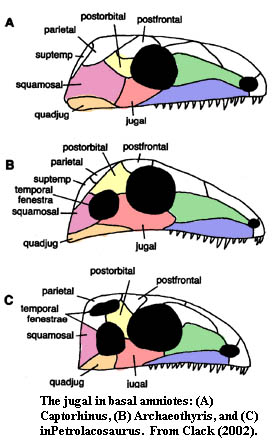
Reptiliomorpha: Here, again, the conservative basal stock maintains the traditional form, but the jugal increasingly tends to separate the maxilla and quadratojugal along the ventral rim of the skull. This is particularly significant because it brings the jugal into contact with the ectopterygoids, giving the jugal at least a peripheral palatal role. As we will see, this has sometimes resulted in a more frank expression of the jugal on the palate. With the introduction of fenestration in the amniote skull, the jugal begins to adopt the triradiate form in which it is commonly found in Mesozoic and Cenozoic tetrapods*.
Anapsida: Many of the more bizarre members of this group, such as the Lanthanosuchidae and Pareiasauria had widely flaring jugals. Turtles seem to have gone almost the other direction. Turtles lack the ectopterygoid bone and have a large medially directed process of the jugal bone. The patterns of skull emargination in the turtles are so varied, that it is difficult to make generalizations, but there are certainly unique arrangements in this group. For example, in the Podcnemoidae, the jugal makes contact with the parietal, the postorbital being very reduced. Gaffney & Meylan (1988). In the Cryptodira, the jugal is a relatively flat bone along the ventral margin of the skull between the maxilla and the quadratojugal.
Eureptilia: Among the eureptiles, the jugal adopts its conventional, triradiate form and becomes quite resistant to change. This (for once) is consistent with the hypothesis we posited in connection with the premaxilla, i.e. that a dermal bone which acquires specialized relationships with different parts of the skull becomes fixed in form. This is not quite correct, but the exception seems to prove the rule. Among Lepidosauriformes, particularly the Squamata, the lower temporal bar tends to be lost, the result being that the jugal is freed from its contact with the quadratojugal and the posterior process is lost. Caldwell & Lee (1997). This liberates the jugal to begin behaving strangely, which it does. So, for example, the jugal of the Anguoidea meets the squamosal above the lower temporal fenestra. Perhaps more typically, as in snakes, the jugal is simply lost.
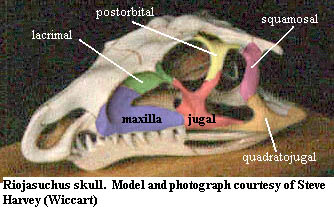 Archosauromorpha: The archosoauromorpha are very conservative indeed, which is scarcely surprising, since the addition of an antorbital fenestra makes the jugal effective tetraradiate. Another fairly constant character of the archosauromorphs is the long scarf joint between the jugal and quadratojugal, as shown in the image of Riojasuchus. A similar joint developes between the jugal and postorbital, giving some degree of mobility to both the posterior and dorsal processes.
Archosauromorpha: The archosoauromorpha are very conservative indeed, which is scarcely surprising, since the addition of an antorbital fenestra makes the jugal effective tetraradiate. Another fairly constant character of the archosauromorphs is the long scarf joint between the jugal and quadratojugal, as shown in the image of Riojasuchus. A similar joint developes between the jugal and postorbital, giving some degree of mobility to both the posterior and dorsal processes.
Continued on Next Page
checked ATW050819