Baphetidae: = Loxommatidae). As normally used, probably a plesion of stem tetrapods, rather than a clade. Here, we stick to the undoubted clade comprised of Baphetes, Loxomma, and Megalocephalus, consistent with [B77].
Introduction: The baphetids, or loxommatids as they were formerly called [ML98], were large aquatic predators of the Carboniferous period. Not only are they included among the very early tetrapods, but they were also among the first of the Carboniferous fossil tetrapods to be found and were originally described in 1850 by William Dawson. They are known mainly from skulls, very little skeletal material. But even from this quite a lot can be deduced. The presence of lateral lines and the long rows of needle-like teeth show that they were fish-eaters. The curious keyhole-shaped orbit indicates the presence either of a gland or, it has been suggested, some sort of electrosensory organ. It may even represent an early form of skull fenestration for jaw muscles. It is not clear whether the four genera assigned to this group are really closely related.
Baphetids have been previously considered primitive temnospondyls and more recently batrachosaurs reptile-like amphibia). It is likely however that they represent one more of a number of early Carboniferous tetrapod radiations. Computer-assisted phylogenetic analyses of a data matrix using characteristics of most of the major groups of terrestrial vertebrates place the Baphetids close to the ancestry of subsequent tetrapod groups. For reference to this, and a number of alternative and previously considered trees, see: Michel Laurin's on-line essay Phylogeny of Terrestrial Vertebrates. It seems from the various alternative trees that the baphetids were cousins of the crassigyrinids, although whether the crassigyrinids evolved from the baphetids as the earlier cladistic trees indicate, see also Panchen ) or the baphetids from the crassigyrinids is not yet certain. (adapted from MAK 930719)
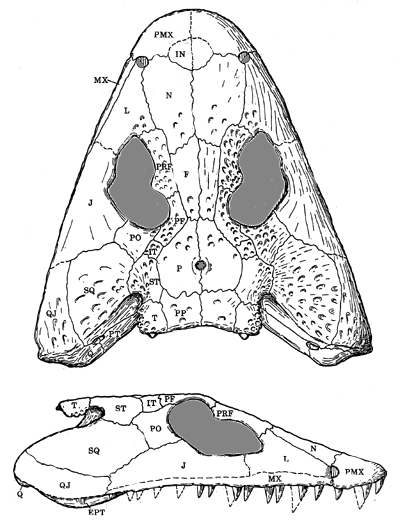
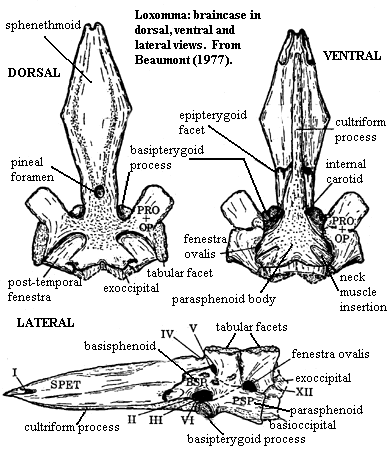 Characters: Supposedly croc-like (post-cranial skeleton essentially unknown until very recently); shallow skull; skull dermal bones similar to Edopoidea [B77]; septomaxilla appears as ornamented, relatively large surface bone [C02]; lateral line grooves present (but somewhat reduced); keyhole-shaped orbits formed by excavation of the lacrimal and prefrontal anterior to the orbit (see figure) and also bordered by jugal [ML98]; antorbital excavation at least as large as orbit [ML98]; strongly embayed spiracular notch, notch at posterolateral border of skull table, opening posteriorly [B77]; small tabulars with posterior process for attachment of mandibular depressors [B77]; orbits at mid-length of skull [B77]; orbits (sensu stricto) circular [B77]; cheek & skull roof sutured; quadrate well behind occiput (as most early tetrapods) [B77]; palatoquadrate probably still present as a unit, although partly formed in cartilage [B77]; exoccipitals do not reach postparietals or continue under skull table (compare Edops) [B77]; supraoccipital absent (not known for all genera) [B77], contra [B00]; stapes proximally broad; palate closed; braincase heavily ossified and extensively attached to skull roofing bones [B77]; opisthotic with two pairs of exoccipital facets (see Megalocephalus) [B77]; nerves IX, X, XI exit between exoccipital facets [B77]; otic capsule with two short, stout columnar processes articulating with exoccipital columns, anterior of which articulates with parietal and supratemporal [B77]; premaxilla form stout buttresses anteriorly on palate [B77]; adsymphysial plate present [ML98]; teeth "labyrinthodont" [C02]; fangs round at base, becoming laterally compressed at crown, with mesial & distal keels [ML98]; parasymphysial plate with at most 2 teeth [ML98]; coronoids have no teeth or denticles; double tooth row on dentary; shows some tendency to consolidation of jaw.
Characters: Supposedly croc-like (post-cranial skeleton essentially unknown until very recently); shallow skull; skull dermal bones similar to Edopoidea [B77]; septomaxilla appears as ornamented, relatively large surface bone [C02]; lateral line grooves present (but somewhat reduced); keyhole-shaped orbits formed by excavation of the lacrimal and prefrontal anterior to the orbit (see figure) and also bordered by jugal [ML98]; antorbital excavation at least as large as orbit [ML98]; strongly embayed spiracular notch, notch at posterolateral border of skull table, opening posteriorly [B77]; small tabulars with posterior process for attachment of mandibular depressors [B77]; orbits at mid-length of skull [B77]; orbits (sensu stricto) circular [B77]; cheek & skull roof sutured; quadrate well behind occiput (as most early tetrapods) [B77]; palatoquadrate probably still present as a unit, although partly formed in cartilage [B77]; exoccipitals do not reach postparietals or continue under skull table (compare Edops) [B77]; supraoccipital absent (not known for all genera) [B77], contra [B00]; stapes proximally broad; palate closed; braincase heavily ossified and extensively attached to skull roofing bones [B77]; opisthotic with two pairs of exoccipital facets (see Megalocephalus) [B77]; nerves IX, X, XI exit between exoccipital facets [B77]; otic capsule with two short, stout columnar processes articulating with exoccipital columns, anterior of which articulates with parietal and supratemporal [B77]; premaxilla form stout buttresses anteriorly on palate [B77]; adsymphysial plate present [ML98]; teeth "labyrinthodont" [C02]; fangs round at base, becoming laterally compressed at crown, with mesial & distal keels [ML98]; parasymphysial plate with at most 2 teeth [ML98]; coronoids have no teeth or denticles; double tooth row on dentary; shows some tendency to consolidation of jaw.
Links: Loxommatidae Baphetidae); Air Breathers of the Coal Period; TREE2000.pdf; Eucritta; AE TREE2000.pdf; Report of Activities 1999: Calder No. 1; Class:Amphibia; Amphibian Skull.
Notes: [1] With the reinterpretation of Ichthyostega and its relatives as aquatic forms, baphetids are good candidates for the spot of first amphibious tetrapod group. If so, baphetids may be a rather important taxon. Both Clack's discovery of Eucritta and recent phylogenetic studies (Ahlberg & Clack (1998)) could support this conclusion. [2] The current candidates for the function of the preorbital excavation are (a) a salt gland, (b) an electrosensory organ; and (c) the pterygoideus muscle. Of the three, the third seems most consistent with the general observation that most serious cranial re-engineering is related to jaw mechanics. This weird adaptation may simply be an early, and very different, type of skull fenestration.
References: Ahlberg & Clack (1998); Beaumont (1977) [B77]; Berman (2000) [B00]; Clack (2002) [C02]; Milner & Lindsay 1988) [ML98]. ATW020820.