Tetrapoda: Overview (2)
 Continued from previous page
Continued from previous page
Palate and Jaw
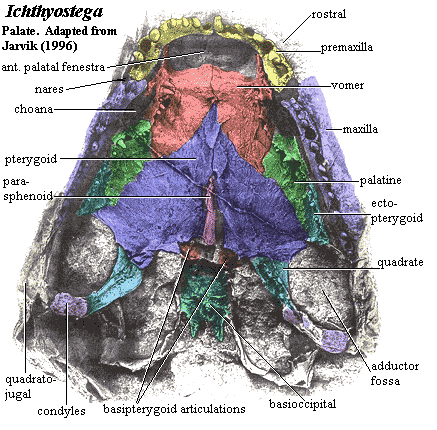
We have hinted that Auntie Stega was not an individual of refined or adventurous tastes; and, indeed, her palate was of a rather conservative type. As mentioned elsewhere, parasymphysial fangs had become increasingly popular in her day. To accommodate these appliances, a number of early tetrapods developed anterior palatal fenestrae [LC93]. Likewise, the vomer exhibited some tendency to expand and develop denticle fields [LC93], and ultimately its own dentition. Certainly, a number of individual adaptations appeared. However, there are no distinctively tetrapod innovations of the palate.
Much the same can be said of the jaw. Auntie Stega was the beneficiary of 20 My of elpistostegalian jaw evolution, which had produced the advanced jaw design discussed in connection with Ventastega. Auntie conserved this legacy but did little more than refine some details. So, for example, the tetrapod splenial developed a medially-directed twist of the ventral margin, exposing the splenial ventrally and mesially [D00]. In other words, the jaw tended toward a sharper keel anteriorly and a more smoothly curved and hydrodynamic ventral surface. Tetrapod jaw adductors were larger, and pulled more vertically. This required reorientation of the articular, but was otherwise compatible with the existing design [D00]. The process of de-ossifying the Meckelian and zippering up the Meckelian fossa continued [AC98].
Axial Skeleton
We have seen that Auntie Stega inherited a well-developed palate and jaw. Sadly, like many a learnèd professional, she was all mouth and no backbone. Granted, the parts were all there. She probably had a rhachitomous vertebral column with 30 or more presacral segments [Co96]. However, the intercentra were paired, not fully fused [Co96]. Zygapophyses were present, but poorly developed [A98] [Co96]. Transverse processes were absent or rudimentary [A98]. The neural arches were well-developed, but the two halves were only partially fused [A98] [Co96]. Auntie Stega did have ribs; and, for the first time, their length generally 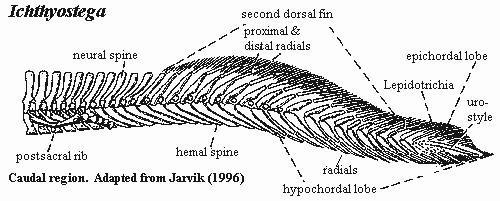 exceeded the combined height of the centrum, neural arch and spine [Co96]. However, it is unlikely that they were much longer, or that they had developed the curvature needed to support the gut against gravity. Auntie did have a single pair of sacral ribs [Co96], and the pelvic girdle (as we will see) was quite substantial. Thus, on the whole, the old girl had all the parts to be a terrestrial type, but they were not yet configured for strolling about.
exceeded the combined height of the centrum, neural arch and spine [Co96]. However, it is unlikely that they were much longer, or that they had developed the curvature needed to support the gut against gravity. Auntie did have a single pair of sacral ribs [Co96], and the pelvic girdle (as we will see) was quite substantial. Thus, on the whole, the old girl had all the parts to be a terrestrial type, but they were not yet configured for strolling about.
Auntie also had all the axial equipment, and properly configured, to be a fish. However, almost all of these accessories were located on the tail. All of the unpaired fins had coalesced into a single structure which wrapped around the end of the tail. This general arrangement is found in many fishes which live in unstable, shallow, fresh water environments, typically with strong seasonal changes, e.g., bichirs, Amia, aruana, and lungfishes [1]. All of these fishes are, like Auntie Stega, predators, and all have additional tetrapod-like behaviors in various combinations: e.g., obligate air-breathing, surface feeding, surface basking, unusual adaptations of the eye, and station-holding using forelimbs. Thus, Auntie and her near-tetrapod kin were not unique in their suite of axial characters. However, for the same reason, these adaptations make very strong  statements about Auntie Stega's environment and mode of life, which we may take up in more detail later.
statements about Auntie Stega's environment and mode of life, which we may take up in more detail later.
Appendicular Skeleton
[2]
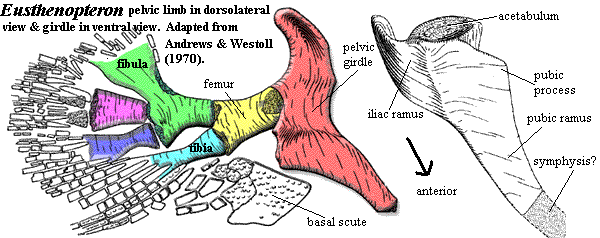
A brief description of the pectoral (forelimb) girdle in Eusthenopteron can be found at this link. Much more basic anatomy of the shoulder girdle in Elpistostegalia can be found here, together with an explanation of the nomenclature used on this page.
As we've mentioned, Auntie's shoulders were almost certainly not part of her head [Co96]. Most of the bones responsible for the head-shoulder connection in Eusthenopteron had been discarded: the supracleithra, posttemporals, and all of the extrascapulars [Co96]. However, the cleithrum retained a postbranchial lamina [Co96], suggesting a strong (or perhaps weak to absent -- [D+94]), residual ligament connection, as well as functional gills. Auntie's cleithrum probably had a rakish posterior slant to it [D+94], perhaps to allow for some remaining degree of coordinated gill-girdle motion through this ligamentous connection [**]. However, in her descendants, the cleithrum assumes a more vertical orientation [**]. Auntie's cleithra, like those of her immediate ancestors, remained fused to the scapulocoracoid [D+94].
 While Auntie was not yet ready to shoulder her way onto land, she was evolving in that general direction. The fossa Tonya was deep, and various scattered scapulocoracoid foramina were beginning to consolidate in that location [**]. The infraglenoid and lateral scapulocoracoid buttresses were becoming more pronounced [**], and the supracoracoid buttress more robust [D+94]; although the coracoid remained a very thin sheet of bone ventrally [D+94]. The glenoid (shoulder joint) itself continued its slow evolutionary movement to increasingly lateral positions [**]. The glenoid cavity was strap-like, suggesting a range of arm motion strongly limited to simple dorsoventral movements [A98] [Co95]. However, the glenoid retained an odd, possibly bimodal curvature hinting at more complex activities -- or perhaps just the ability to fold her arms to the side when not needed [**]. Auntie's clavicles had a broad ventral plate and narrow ascending processes [C97], and they articulated with an elongate, diamond-shaped interclavicle.
While Auntie was not yet ready to shoulder her way onto land, she was evolving in that general direction. The fossa Tonya was deep, and various scattered scapulocoracoid foramina were beginning to consolidate in that location [**]. The infraglenoid and lateral scapulocoracoid buttresses were becoming more pronounced [**], and the supracoracoid buttress more robust [D+94]; although the coracoid remained a very thin sheet of bone ventrally [D+94]. The glenoid (shoulder joint) itself continued its slow evolutionary movement to increasingly lateral positions [**]. The glenoid cavity was strap-like, suggesting a range of arm motion strongly limited to simple dorsoventral movements [A98] [Co95]. However, the glenoid retained an odd, possibly bimodal curvature hinting at more complex activities -- or perhaps just the ability to fold her arms to the side when not needed [**]. Auntie's clavicles had a broad ventral plate and narrow ascending processes [C97], and they articulated with an elongate, diamond-shaped interclavicle.
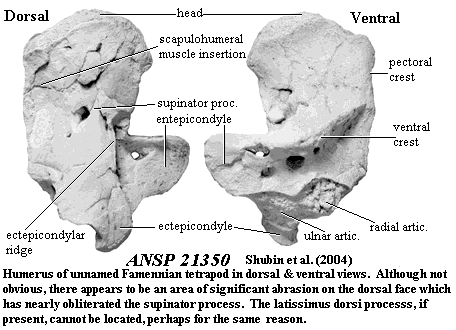
Auntie Stega's humerus was flat and L-shaped [A98] [Co96]. It had become "a broad platform for the insertion of extensor and flexor muscles that [was] less mobile than the humeri of Eusthenopteron and other basal tetrapodomorphs" [S+04]. The anterior margin was sharp, bearing a strong pectoral process [A98]. The supinator ridge or process also lay close to anterior margin [A98]. The entepicondyle was very large and probably rectangular [Co96] The tall, slender ectepicondyle arose from a strong ectepicondylar ridge and extended well below the distal end of the humeral shaft [Co96] [A98] [S+04]. Where the ectepicondyle and entepicondyle nearly meet, we see a deep recess [S+04]. In her descendants, a latissimus dorsi ridge was present there, generally continuous with the ectepicondylar ridge [S+04]. The ventral, or "oblique" ridge remained a dominant feature of the ventral surface, pierced by large foramina. This ridge ran nearly perpendicular to the long axis of the humeral shaft and was confluent with the posteromedial rim of the entepicondylar process [S+04]. The epipodial facets i.e. the articulations for ulna and radius) were probably separated [Co96].
 The epipodials themselves remained quite primitive. Auntie's radius was distally quite flat and broad [Co96]. The ulna was short and lacked an olecranon process [Co96]. The intermedium articulated only proximally and distally -- that is, it did not articulate with other wrist bones [Co+02]. Auntie had no distal carpals [Co+02]. In fact there was little specialization of the wrist or ankle bones, although they were soon to develop [LCo95]. On the other hand, so to speak, Auntie's had fingers -- in fact she probably had 6-8 fingers on each hand.
The epipodials themselves remained quite primitive. Auntie's radius was distally quite flat and broad [Co96]. The ulna was short and lacked an olecranon process [Co96]. The intermedium articulated only proximally and distally -- that is, it did not articulate with other wrist bones [Co+02]. Auntie had no distal carpals [Co+02]. In fact there was little specialization of the wrist or ankle bones, although they were soon to develop [LCo95]. On the other hand, so to speak, Auntie's had fingers -- in fact she probably had 6-8 fingers on each hand.
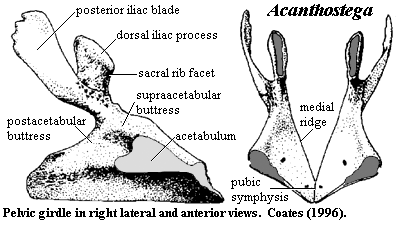
Generally speaking, Auntie Stega's arms weren't all that different from the forelimbs of Eusthenopteron, except that they bore fingers instead of fin rays. However, Auntie had great legs. Compare the pelvis of Acanthostega with the pelvic girdle of Eusthenopteron. It's difficult even to recognize them as the same structure. Each side of Auntie's pelvic girdle was still composed of a single bone [Co96]. To that extent, the structure was still fish-like. But the two halves of the pelvic girdle were sutured together all along the ventral margin to form a single, smoothly curved ventral surface. Dorsally, the hips were attached to the spine by small sacral area involving a specialized sacral rib [Co96]. The pelvic girdle had two iliac processes (albeit oddly constructed), an anteroventral pubic symphysis, a laterally-facing acetabulum, and a supraacetabular buttress. Auntie's hips were, therefore definitively tetrapod hips -- not fundamentally different from 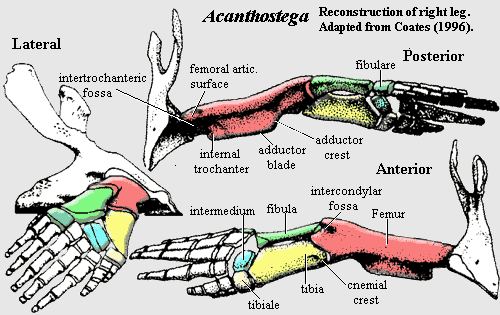 the hips of an early archosaur, for example.
the hips of an early archosaur, for example.
Auntie's femur looked strikingly like the femur of a very early synapsid [Co96]. To some degree this is coincidence, since the legs of many of Auntie's more immediate descendants look less like those of a "pelycosaur." Nevertheless, it is indisputably a tetrapod leg. None of the limb elements is measurably twisted [Co96]. The femoral adductor blade and crest are well-developed [A98] [Co96]. A rugose fourth trochanter is present and restricted to adductor blade [Co96]. Between the distal condyles of the femur we can clearly see an intercondylar fossa, presumably the precursor of the popliteal fossa [A98]. The epipodials are parallel, and both articulate with identifiable tarsal elements. The tibia has an L-shaped distal articular surface (since it articulates with two tarsals) and a cnemial crest [Co96]. At least some of the tarsal elements may articulate laterally as well as proximodistally [Co96], although that development may post-date Auntie. Typically for an early tetrapod, the fibulare and intermedium are large and each articulates with several digits. Finally, Auntie Stega's toes were much longer than her fingers.
Auntie Stega's Condition
This concludes our head-to-toe survey of the Mother of All Tetrapods. As usual, we have had to deduce her characteristics by triangulating through her immediate neighbors in phylospace. However, as it turns out, those characteristics are consistent enough that we can draw a very detailed picture of the old girl. What we see is a very mixed bag: a few startling innovations, such as the otic region of the brain, a few parts that hardly changed at all, like the palate -- but mostly the continuous, if slightly irregular, ticking of time and evolution, constantly remodeling anatomy in small increments.
References: Ahlberg 1995) [A95]; Ahlberg (1998) [A98]; Ahlberg & Clack (1998) [AC98]; Ahlberg & Luksevics (2004)[AL04]; Ahlberg, et al. (1996) [A+96]; Clack 1989) [C89]; Clack (1994) [C94]; Clack 1997) [C97]; Clack (1998a) [C98]; Clack (2002) [C02]; Clack 2002a) [C02a]; Clack, et al. 2003) [C+03]; Coates (1996) [Co96]; Coates & Clack (1991) [CoC91]; Coates, et al. (2002) [Co+02]; Daeschler (2000) [D00]; Daeschler, et al. (1994) [D+94]; Jarvik 1996) [J96]; Lebedev & Clack (1993) [LC93]; Lebedev & Coates (1995) [LCo95]; Manley & Clack (2004) [MC04]; Shubin, et al. (2004) [S+04].
ATW050530. Text public domain. No rights reserved.
[1] For some reason, the early tetrapods are frequently compared to a totally different ecomorph, the short-range ambush predators exemplified by pike. We have discussed elsewhere the many reasons why these body forms are mechanically dissimilar.
[2] In the process of working up materials on the elpistostegalian shoulder girdle, we noticed some minor patterns that weren't expressly stated by others in the articles we reviewed. To avoid confusing these speculations with reliable information, we've marked them with a double asterisk [**].

{kind=link}