 Demorphing the Tetrapodomorpha: 1. Myths and Mything Links
Demorphing the Tetrapodomorpha: 1. Myths and Mything Links| Sarcopterygii | ||
| The Vertebrates | Tetrapodomorpha |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
|
Contents
Overview |
Demorphing the Tetrapodomorpha: 1. Myths and Mything LinksTwo generations ago Prof. Alfred S. Romer explained the emergence of tetrapods as a gradual adaptation of osteolepiform fishes moving between waterholes during "the periodic droughts of the Devonian." Much fun has been poked at this well-known "process explanation" or "scenario" approach to evolutionary theory. The human mind is good at making stories to explain things. That's what mythology is all about. In the Middle Devonian sea there were various rhipidistian fishes. By the Mississippian, some of their ancestors lived on land. The mind then constructs, unbidden, a suitable shorthand vision of the process, the nature of which may depend on one's experience, culture, and degree of interest. Every ancient civilization used myth as a tool of science. We're not that different.
We know, of course, that it was nothing like that. There was no story line or teleology which drove the business. But science is always in tension between what we know and the way we learn. We learn by patterns, scenarios, processes, analogies, pictures and stories. We extrapolate and interpolate from them to specific cases. The burden of remembering all that we must know without these tools would be too great. Yet, the facts don't necessarily arise from those processes, or any others. Particularly in paleontology, our data are the scattered products of genetic chance, preservational artifact, and observational selection. The job at hand is to reconcile these elements as best we may. Thus a scenario or process explanation which reasonably accounts for what we know at a particular point in time is not a bad thing, so long as we understand its hybrid nature. It is not historical fact, but may allow us to describe and relate historical facts too complex to confront directly. It is not a testable hypothesis, but it may lead to the formulation of testable theories. It is not a scientific cause of observable effects, but more akin to the legal concept of proximate cause -- factors which tilt the scales of chance in some direction. Ultimately, these sorts of conceptual constructs are not really intended as explanations of anything. Rather, they are what most paleontology has always been about: describing what we see so that it can be related to other observations. If we are careful not to take ourselves too seriously, the scenario, the process explanation, and the myth are useful and appropriate bridges between our learning and our knowledge.
 In that spirit, we will attempt to look at the demorphing of tetrapodomorphs. How -- and a little bit of why -- one group of rhipidistian fishes evolved the tetrapod condition. For those with the time and inclination, Dr. Jenny Clack's 2002) recent book is strongly recommended as a more complete, and certainly much more authoritative, treatment of the subject.
In that spirit, we will attempt to look at the demorphing of tetrapodomorphs. How -- and a little bit of why -- one group of rhipidistian fishes evolved the tetrapod condition. For those with the time and inclination, Dr. Jenny Clack's 2002) recent book is strongly recommended as a more complete, and certainly much more authoritative, treatment of the subject.
What we will find is that at least four distinct lineages of sarcopterygian fishes developed some tetrapod-like adaptations in the Late Devonian and earliest Carboniferous: the lungfishes, rhizodonts, tristichopterids and elpistostegalians (we will not cover the lungfishes, except incidentally.) Not surprisingly, there is a high degree of homoplasy or "convergent evolution." The overwhelming consensus view is that we evolved from the elpistostegalians. However, some tetrapod adaptations are not known in non-tetrapod Elpistostegalia, but are known in some their competitors. Thus, for example, some rhizodonts appear to have had digits on their forefins like tetrapods, although the advanced elpistostegalian, Panderichthys, had none. Lungfish obliterated the division between the anterior and posterior halves of the braincase (the intracranial joint) as did tetrapods. Panderichthys retained the same sharp division of the brain as its eldest rhipidistian ancestor. So, it is still possible -- if rather unlikely -- that the tetrapods are polyphyletic, as Erik Jarvik thought, or that they derived from outside the Elpistostegalia. Coates et al. (2002).
One currently widespread belief, often associated with the names of Dr. Clack and Dr. Per Ahlberg, is that very few of the "tetrapod" adaptations are actually adaptations for life on land. Many seem to be design changes in response to selective pressures for more efficient aquatic life. In fact, Dr. Clack has argued that Acanthostega, which plainly is a tetrapod, seldom, if ever, ventured on land. Thus the tetrapod condition is a suite of aquatic specializations which just fortuitously allowed life on land. We will offer some reasons why we believe this position overstates the case.
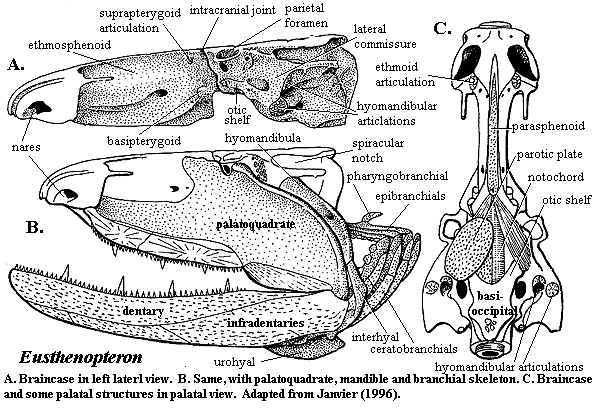
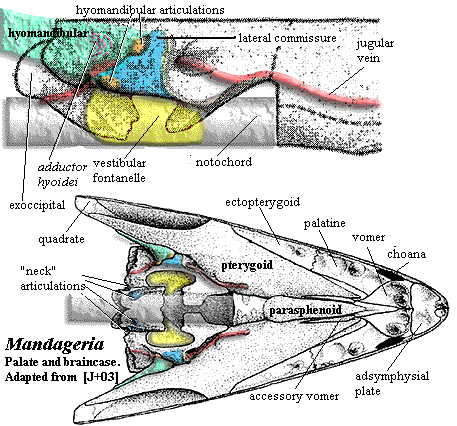
Finally, for historical reasons, almost all descriptions of tetrapodomorphs refer frequently to Eusthenopteron. The exhaustive studies of Jarvik, and of Andrews and Westoll (1970) on many complete specimens of this tristichopterid from the Famennian Miguasha formation of Quebec have ensured that it will be one of the best-known vertebrates of the entire Paleozoic. Our discussion will be no different, although we will also look frequently to the remarkable work of Dr. Zerina Johanson and her colleagues on the much more specialized tristichopterid Mandageria. Johanson & Ahlberg (1997), Johanson et al. (2003). It may be worth reviewing the following detailed figures of these fishes found elsewhere on this site:
GO TO PART 2. THE DERMAL SKULL
or referring to them as needed. ATW031026.

The Tetrapodomorpha are defined as Londoners > lungfish. Basally, they differ little from the basic Rhipidistian pattern. The synapomorphies of the group, as determined by Cloutier & Ahlberg (1996), are relatively small matters of head and hand. In the head, the pineal foramen is open. The parasymphysial tooth whorl is lost, and the vomers meet on the midline of the palate. Something is clearly going on with the nares, but there is much disagreement about exactly what.
These are matters of detail, but they all suggest that a critical evolutionary corner had been turned. As we have noted many times, vertebrate evolution is very frequently driven by jaws or their functional equivalents. In this department, the early osteichthyans, including the Rhipidistian stem group, faced a common constraint. The anterior skull was quite short, limiting the size of the jaws. Lengthening the jaw required some solution to a difficult design problem: how to sufficiently brace the upper jaw against mechanical stress when it moved out from under protection of the braincase and skull table. Broadly speaking, each of the three rhipidistian clades represents a different solution to the problem. The Porolepiformes made the anterior skull broader, substituting additional width for length. The Dipnoi consolidated the small bones of the rostrum into a rigid rostral shield, while strengthening and simplifying the structure of the jaw. The Tetrapodomorphs took a more difficult, but more elegant approach. They lengthened the skull table, creating a long, complex arch of elongate medial bones from snout to occiput. Eventually, this approach was to have profound implications for the structure of the brain, among many other things. But, in the Mid-Devonian, it was reflected only in small things: parietals with enough stability to open an immobile foramen to the pineal, vomers that met in the midline helped anchor one end of the incipient arch, reorganization of the nares which may have eliminated a lateral point of weakness.
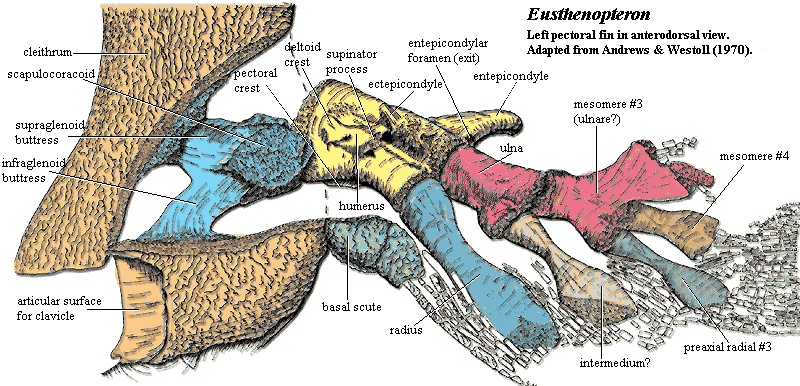
Even more significant may be the changes in the pectoral limb. The head of the tetrapodomorph humerus is convex, rather than concave, so that the fin rotates in the glenoid fossa of the scapulocoracoid. The difference in basal forms was slight, both surfaces being relatively flat. However, the difference in the long term was probably critical to the development of the tetrapod forelimb. For weight-bearing, terrestrial locomotion, it makes much better engineering sense to have the concave element inside the body. In that way, the sides of the joint, which experience great stress, can be stabilized by non-moving body elements. If the humeral head were concave, the humerus would have to be much more massive to buttress the concavity against the lateral forces experienced by the walls of the socket.
Just as important, the limb itself was uniserial: built off a single metapterygial axis and with fin radials extended only from the post-axial side of the fin. This may bespeak an important change in locomotor style. Most fishes do not use their forelimbs for motive power. That's the function of the tail. The pectoral fins are used for braking, turning, and attitude control. Such fins need to be very flexible and designed so that they can trim the animal's hydrodynamic profile in any arbitrary way. Thus, they tend to be relatively flat and symmetrical, frequently with considerable surface area. The basic tetrapodomorph pattern is asymmetrical and intrinsically narrower. This makes a very efficient, high aspect ratio hydrofoil for cruising, but gives poor mobility at low speeds. On the other hand -- so to speak -- the design opens up many new possibilities for powered locomotion. These range from bottom-walking to whatever it was the plesiosaurs did (see discussion) to, eventually, walking on land. ATW031026.
 The Rhizing Tide
The Rhizing TideThe rhizodonts were predatory fishes. Often huge, they may have reached sizes of six meters or more. Not all were quite so massive, of course. The pictured reconstruction of Gooloogongia was based on a specimen measuring a mere 90 cm. The rhizodonts are first known from the Middle Devonian. They flourished worldwide in the Mississippian, but became extinct before the end of the Carboniferous. Although much progress has been made in recent years, rhizodonts are still rather poorly known. Our knowledge of them suffers not only from the usual rarity of top predators, but from their very loosely constructed skull which tended to fall apart rapidly after death.
Rhizodonts, like various other top predators, appear to have been shaped solely by an imperative to cram as much down their throats as possible in a minimum amount of time. Their mouths are large. The teeth are blade-like, strongly held, and reinforced internally with bridges of bone between dentine folds. The dentition included huge dentary tusks. The jaws are not those of the typical long-snouted fish-eater. They are broad and C-shaped, like the jaws of their contemporaries, the Early Carboniferous porolepiforms. The gape of the jaws is impressive, and was further enlarged by the flexible skull and strongly kinetic intracranial joint.
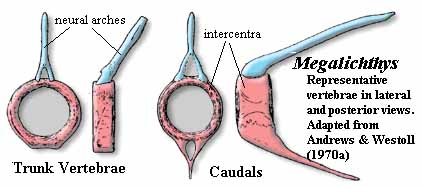
As in other tetrapodomorph lineages, the axial skeleton is rather variable. In Strepsodus, the vertebral centra were ring-like, as in Megalichthys see image); but in Rhizodus, the centra were weakly ossified and remain poorly known. All of the fins, except the pectorals, were small and placed posteriorly. In fact, the anal and posterior dorsal fins were functionally part of the caudal fin. This is a pattern we will see in the derived members of all rhipidistian groups. The dorsal fins were supported by a basal element articulating with one vertebra each. This base flared distally, and bore four proximal radials, each with one or two rows of distal radials. The tail fin tended to be diamond shaped and diphyceral -- again consistent with the common tendency of all tetrapodomorph groups.
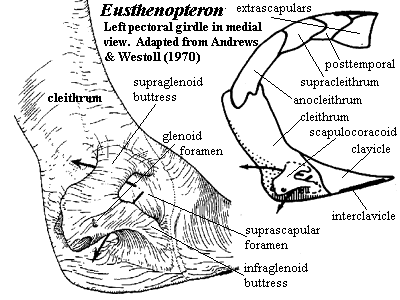
 The forefins are built on a very different model. They are massively braced, comparatively rigid and powerfully supplied with muscles. In fact, even in such a large powerful carnivore, the forefins seem significantly over-engineered. As with all tetrapodomorphs, the biggest component of the shoulder girdle is the cleithrum (see images below), a dermal bone connected to the skull by a chain of smaller bones (see inset in Eusthenopteron image). In rhizodonts the cleithrum is not only massive in itself, but located quite far anteriorly and closely integrated with the skull through soft-tissue connections. It is deeply inset to accommodate large muscles, particularly the ventral pectoralis. The cleithrum is also braced with a double articulation on the clavicle ventrally. The tetrapodomorph scapulocoracoid is typically a relatively slight, tripodal structure on the inner surface of the cleithrum. In rhizodonts, the scapulocoracoid was a massive block of endochondral bone.
The forefins are built on a very different model. They are massively braced, comparatively rigid and powerfully supplied with muscles. In fact, even in such a large powerful carnivore, the forefins seem significantly over-engineered. As with all tetrapodomorphs, the biggest component of the shoulder girdle is the cleithrum (see images below), a dermal bone connected to the skull by a chain of smaller bones (see inset in Eusthenopteron image). In rhizodonts the cleithrum is not only massive in itself, but located quite far anteriorly and closely integrated with the skull through soft-tissue connections. It is deeply inset to accommodate large muscles, particularly the ventral pectoralis. The cleithrum is also braced with a double articulation on the clavicle ventrally. The tetrapodomorph scapulocoracoid is typically a relatively slight, tripodal structure on the inner surface of the cleithrum. In rhizodonts, the scapulocoracoid was a massive block of endochondral bone.
The limb is not only notable for its supports, but its structure. In several ways it is significantly more like the tetrapod forelimb than, for example, the fore-fin of Panderichthys. The humerus has a distinct, offset head, like tetrapod, and was apparently capable of a very wide range of motion. Unlike any other fishes, rhizodonts also possessed what can only be called digits, as the image shows. The fin rays (lepidotrichia) sprouting from the ends are also unique in having a long, stiff unsegmented basal area. In short, considering the pectoral limb alone, rhizodonts would appear to be much closer to the tetrapods than any other sarcopterygian group. Coates et al. (2002).
There is increasing evidence that all tetrapodomorphs, including the earliest tetrapods, were aquatic. If so, one has to wonder if the rhizodonts were not, after all, the first on the beach. ATW031028.
Image: Gooloogongia loomesi from the Age of Fishes Museum site. Note that the pectoral fin as reconstructed in the figure is much more ventrally placed and directed than may have been the case in Rhizodus. Jeffery's reconstruction of Strepsodus (after Andrews) shows a somewhat intermediate condition.
Tetrapodomorpha: Tetrapoda > Dipnoi. Kenichthys.
Range: from at least the Early Devonian
Phylogeny: Rhipidistia : Dipnomorpha + * : Rhizodontiformes + Osteolepiformes
Characters: only anterior nares present [CA96$] [2]; pineal foramen open [CA96$]; supratemporal present [VS91]; vomers articulate on midline [CA96$]; paired intervomerine pits absent [CA96$]; dentary without parasymphysial tooth whorl [CA96$]; lateral extrascapulars overlap median [VS91] (contra [CA96]: plesiomorphic for Sarcopterygii); median gular present [VS91] (contra [CA96]: rhipidistian synapomorphy); short, uniserial paired fins [S95] [1]; humerus with convex head articulating with glenoid fossa [VS91] [CA96$]; humerus with deltoid & supinator processes [CA96$].
Notes: [1] According to Shubin [S95] these characters are primitive for gnathostomes. [2] Cloutier & Ahlberg [CA96] assert that porolepiforms have a palatal opening homologous to the choana.
Links: Comparative Anatomy Topic 3 - Fishes a few basic facts).
References: Coates et al. (2002) [C+02], Cloutier & Ahlberg (1996) [CA96], Vorobyeva & Schultze (1991) [VS91]. ATW031004.
 Rhizodontiformes: = Rhizodontida) Barameda, Gooloogongia, Rhizodus, Sauripteris, Screbinodus, Strepsodus (includes Archichthys).
Rhizodontiformes: = Rhizodontida) Barameda, Gooloogongia, Rhizodus, Sauripteris, Screbinodus, Strepsodus (includes Archichthys).
Range: Givetian to Pennsylvanian, mostly Mississippian.
Phylogeny: Tetrapodomorpha : Osteolepiformes + *.
Characters: many very large (jaw > 100 cm); dermal bones thin & loosely attached [J96]; intracranial joint present [CA96]; intracranial joint allowed very large gape [L95]; postparietal elongate, very large [J96]; lateral extrascapulars triangular & approach closely on midline [JA97]; extrascapulars very large [J96]; parasphenoid large [J96]; choanae believed to be absent; lower jaw C-shaped in cross section - hollow tube lingually incomplete [1]; prearticular with large ascending process; teeth polyplocodont, with bone bridges between folds; upper jaw with marginal row of small teeth on maxilla & premaxilla, medium-sized fangs on ectopterygoid and palatine & large tusks on vomers & premaxillae; premaxilla may have posterior palatal process with large median tooth (fang?) [JA97]; vomers lack posterior process [JA97]; parasymphysial fang pair present [JA97]; parasymphysial plate small, not contacting coronoid (Notorhizodon) [J96] [JA97]; lower teeth include marginal teeth on dentary, fangs on three coronoids & huge tusk at dentary symphysis [CA96]; some species with laterally compressed blade-like teeth [L95]; trunk elongate; vertebrae thinly ossified in Rhizodus[AW70a]; centra ring-like, thick, strongly ossified amphicoelous and elongate in Strepsodus [AW70a]; zygapophyses, parapophyses & ribs absent [AW70a]; neural arches simple [AW70a]; all fins (except pectorals) much reduced and placed posteriorly (anal & 2nd dorsal fins were functionally part of the caudal fin); dorsals supported by element articulating with one vertebra, thin and straight proximally, flaring distally, and bearing 4 proximal radials, with 1-2 rows of distal radials [AW70a] [6]; anal fin probably same as dorsals [AW70a]; caudal fin tended to be diamond shaped and diphyceral; cleithrum very large, with triangular to ovate ventral lamina [AW70a]; paired fins with approximately 5 mesomeres (segments) [C+02]; cleithrum dorsal process elongate and rotated with respect to ventral lamina so that outer surface faces anterolaterally [AW70a] [3]; cleithrum dorsal lamina with unornamented anterior margin presumably for gill lining membrane [AW70a]; unornamented area expanded dorsally into anterodorsal process, possibly for membranes channeling water leaving gills [AW70a]; cleithrum with dual articulation for clavicle as shown in figure [AW70a] [J96]; cleithrum with "depressed lamina" [CA96$]; cleithrum ventral lamina, medial face with strong posterior ridge marking limit of pectoralis attachment as in Eusthenopteron [AW70a]; cleithrum with widely expanded ventral lamina [J96]; cleithrum medial face with structures shown in figure [AW70a] [4]; clavicle located far anteriorly [AW70a]; clavicle lacks anterior muscle attachment area (compare Eusthenopteron) [AW70a]; clavicle with strong, hollow,  posterodorsal spine for dorsal articulation with cleithrum [AW70a] [5]; interclavicle long, and narrow [AW70a]; pectoral fins strongly enlarged [C+02]; scapulocoracoid apparently not tripodal; well developed pectoral endoskeleton surrounded by robust, largely unsegmented lepidotrichia [CA96$] [J96]; and covered in deeply overlapping scales (creates broad paddle) [AW70a] [L95]; forefins generally broad & fan-like [C02]; pectoral basal scutes unknown and probably absent [AW70a]; glenoid fossa deep & circular, allowing good all-round movement of the paddle [2]; humerus distinct, with caput humeri and a long entepicondylar process (but no other mesomeres have postaxial process) [AW70a]; humeral head offset by neck, rounded and bulbous [JA97]; humerus with well defined ectepicondyle, supinator & deltoid processes [AW70a]; may be ectepicondylar foramen between supinator & deltoid processes, and also a latissimus dorsi process [AW70a]; entepicondylar foramen also present [AW70a]; humerus distal articulations confluent, with convex radial articulation, flat ulnar articulation [AW70a]; 3rd mesomere (centrale?) has no postaxial process but articulates with elements of a possible digital arch as in tetrapods segmented radials) [AW70a] [J96] (see image); pelvic girdle bar-like, with unossified symphysis [C+02]; acetabulum flanked by pubic & iliac processes [C+02]; scales round, thin & without cosmine [CA96] [J96]; scales ornamented with low ridges and a central boss on the inner surface [AW70a]; lateral line system elaborated on skull & pectoral girdle.
posterodorsal spine for dorsal articulation with cleithrum [AW70a] [5]; interclavicle long, and narrow [AW70a]; pectoral fins strongly enlarged [C+02]; scapulocoracoid apparently not tripodal; well developed pectoral endoskeleton surrounded by robust, largely unsegmented lepidotrichia [CA96$] [J96]; and covered in deeply overlapping scales (creates broad paddle) [AW70a] [L95]; forefins generally broad & fan-like [C02]; pectoral basal scutes unknown and probably absent [AW70a]; glenoid fossa deep & circular, allowing good all-round movement of the paddle [2]; humerus distinct, with caput humeri and a long entepicondylar process (but no other mesomeres have postaxial process) [AW70a]; humeral head offset by neck, rounded and bulbous [JA97]; humerus with well defined ectepicondyle, supinator & deltoid processes [AW70a]; may be ectepicondylar foramen between supinator & deltoid processes, and also a latissimus dorsi process [AW70a]; entepicondylar foramen also present [AW70a]; humerus distal articulations confluent, with convex radial articulation, flat ulnar articulation [AW70a]; 3rd mesomere (centrale?) has no postaxial process but articulates with elements of a possible digital arch as in tetrapods segmented radials) [AW70a] [J96] (see image); pelvic girdle bar-like, with unossified symphysis [C+02]; acetabulum flanked by pubic & iliac processes [C+02]; scales round, thin & without cosmine [CA96] [J96]; scales ornamented with low ridges and a central boss on the inner surface [AW70a]; lateral line system elaborated on skull & pectoral girdle.
Images: The fossil skull of Barameda has been labeled by analogy to Eusthenopteron. Long [L95] uses the more traditional actinopterygian nomenclature.
Note: [1] On his site, from which some of the description above is derived, Jeffery notes that, "on biting, the left and right mandibles would rotate inwards towards each other. This may be a kinetic mechanism to dig the marginal teeth more deeply into the prey, to help grip slippery or struggling items. The upper jaw also appears to have been quite kinetic, with loosely fitting bones surrounding a cartilaginous braincase." [2] Jeffrey also states that "this suggests that rhizodonts could generate powerful thrashing and dragging movements, which could be used to manipulate and dispatch prey." [3] Note how the rhizodont cleithrum converges on the  morphology of the tetrapod scapulocoracoid. This may be even more true of other rhizodonts, e.g., Strepsodus. [4] The cleithrum is flattened during fossilization. This appears to account for the morphological difference between the reconstruction and the photograph of this bone. [5] In Megalichthys, the dorsal spine of the clavicle passes medial to the cleithrum, while the ventral overlap is lateral. This is reversed in Rhizodus [AW70a]. These are actually rather complex curves, and the case may be more complicated. [6] the arrangement looks suspiciously digit-like and similar to the 3rd mesomere of the pectoral fin.
morphology of the tetrapod scapulocoracoid. This may be even more true of other rhizodonts, e.g., Strepsodus. [4] The cleithrum is flattened during fossilization. This appears to account for the morphological difference between the reconstruction and the photograph of this bone. [5] In Megalichthys, the dorsal spine of the clavicle passes medial to the cleithrum, while the ventral overlap is lateral. This is reversed in Rhizodus [AW70a]. These are actually rather complex curves, and the case may be more complicated. [6] the arrangement looks suspiciously digit-like and similar to the 3rd mesomere of the pectoral fin.
Links: Rhizodontida Mikko's Phylogeny); Rhizodonts - Big Dead Fish Best on the Web); First East Gondwanan record of Strepsodus (Sarcopterygii, ....
References: Andrews & Westoll (1970a) [AW70a], Clack 2002) [C02], Cloutier & Ahlberg (1996) [CA96], Coates et al. (2002) [C+02], Janvier 1996) [J96], Johanson & Ahlberg (1997) [JA97]; Long 1995) [L95]. ATW031009.
checked ATW060320
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}