
| Tetrapoda | ||
| The Vertebrates | Ichthyostega |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Sarcopterygii
│
└─Tetrapoda
├─Ichthyostega
└─┬─Hynerpeton
└─┬─Tulerpeton
└─┬─Whatcheeriidae
└─┬─Crassigyrinus
└─┬─Colosteidae
│ └─Greererpeton
└─┬─Baphetidae
└─Tetrapoda*
├─Temnospondyli
└─┬─Lepospondyli
└─Reptiliomorpha
|
Contents
Overview |
| ...
Þá gén sylf cyning Féond gefyldan / -- ferh ellen wræc -- Ond hí hyne þá bégen / ábroten hæfdon, Þegn æt ðearfe. / Þæt ðám þéodne wæs Síðas sigehwíle / sylfes daédum, |
... Once more the old king, Drew deep on his Will, drew out the death dagger, Steel sharpened by battle, on iron mail lashed. Struck Lord of the Wederas! Knife gutting dragon! Cut demon down dead - for courage breeds strength -- And so both together created destruction, As brothers in arms, as each man should stand For the other in troubles. That was the Chief's way, His high road to triumph, to the last of his Deeds, Of his work in this world. ... |
| Beowulf XXXVII, ll 2702-2711 | Transl. ATW050806 |
Below is the usual compressed summary of the characteristics of Ichthyostega. In preparing this summary, we have done our usual incomplete survey of the descriptive literature; and, like a blackbird, we have probably picked up shiny facts more for their intrinsic attraction than for their significance. However, in the case of Ichthyostega we've made an important exception. We have read, but have included nothing from, Jarvik 1996). Jarvik (1996) is the only monograph on Ichthyostega, a very important early tetrapod; and it was written by a scientist whom we have frequently praised for his work on Eusthenopteron and other osteolepiform fishes. The reason for this deliberate omission is simply that Jarvik (1996) is not a reliable source. Here we will first justify that statement and then examine some of the historical background of the paper, with an eye toward some of the odd effects this state of affairs has had on the study of early tetrapods.
The problems with Jarvik's study of Ichthyostega seem to be well known in the small community of early tetrapod workers. A brief, and by no means exhaustive, summary of the literature reveals the following. Säve-Söderberg originally named and identified about five different species of Ichthyostega, together with a supposed sister genus, Ichthyostegopsis. However, "Jarvik never really attempted to recognize or verify the presence of the different species" and simply lumped all of the material under I. stensioei without significant discussion of either morphology or geological setting [B05]. The dermal bones of the skull were described using a rather bizarre and certainly unique system of nomenclature drawn in part from supposed resemblances to the skulls of humans and frogs [J96]. Ahlberg & Clack [ACl98] concluded that Jarvik had misinterpreted the medial face of the inner jaw in various significant details, including the course of the oral sensory canal. In the palate, he ignored the presence of an ectopterygoid [B05].
Although Jarvik was the custodian of most known remains of Ichthyostega for fifty years, he never really attempted to explore, compare or understand its strange braincase [B05]. What little work he did on the braincase included misidentification of both the stapes and inner ear [Cl02a] [Cl+03]. Jarvik's description of the spine was at least partially "idealized" and inaccurate, and he misidentified both the sacral rib and its attachment point on the pelvis [Co96]. Blom bluntly states that "the braincase and the vertebral column, still remain to be explored." [B05] Jarvik's reconstruction of the pectoral girdle exaggerates the post-branchial lamina and includes a large supraglenoid process which appears to be an artifact [LCo95]. He plainly found 7-8 digits, but insisted that Ichthyostega had only five. Other, vaguer, grumblings in the literature suggest that this by no means exhausts the list of problem areas.
 Jarvik was 89 years old when the article finally went to press. Not all of us will reach the age of 89, and few of us indeed are likely to be inclined or inspired to write a massive monograph on a unique tetrapod at that age -- even assuming we were capable of doing so at some earlier point. So, perhaps Jarvik's work does not reflect failure. It only suggests lack of complete success at a task which almost no one else would even have attempted in the first place. But, how did it happen that Jarvik took on this immense challenge at such an unlikely point in his career? The story has some odd resonances with the epic of another aging Scandinavian who struggled to lift a curse and obtain the treasure of another unique creature -- the saga of Beowulf and the Fire Dragon with which we began this section.
Jarvik was 89 years old when the article finally went to press. Not all of us will reach the age of 89, and few of us indeed are likely to be inclined or inspired to write a massive monograph on a unique tetrapod at that age -- even assuming we were capable of doing so at some earlier point. So, perhaps Jarvik's work does not reflect failure. It only suggests lack of complete success at a task which almost no one else would even have attempted in the first place. But, how did it happen that Jarvik took on this immense challenge at such an unlikely point in his career? The story has some odd resonances with the epic of another aging Scandinavian who struggled to lift a curse and obtain the treasure of another unique creature -- the saga of Beowulf and the Fire Dragon with which we began this section.
In the saga, the aging Beowulf attacks the red dragon. He is helped only by a very young warrior named Wiglaf. Beowulf's sword breaks, and he is terribly wounded. But Wiglaf comes in under Beowulf's shield (his own having been reduced to charcoal) and delivers a huge blow which snuffs the dragon's flame. Beowulf seizes the chance to pull out his small wællseaxe -- normally used for routine camp chores, such as butchering game and dispatching wounded enemies. With a last, huge effort, Beowulf disembowels [1] the dragon; but he dies a few minutes later, after a suitably dramatic speech to Wiglaf. Jarvik's story is more complex and multi-generational -- actually more like a traditional Norse epic than Beowulf in some ways -- and he was both Wiglaf and Beowulf at various points.
The first Late Devonian tetrapods were found in 1929 in rocks dating from the upper Famennian stage of East Greenland (on the north slope of Celsius Berg) by Oskar Kulling of Sweden. He sent his material to Erik Stensiö 1891-1984) at the Swedish Museum, who was intrigued, recognizing them to be scales of a fishlike vertebrate of uncertain identity.Stensiö had two serious students at the time: Erik Jarvik and an even younger man (who may have been an undergraduate at the time), Gunnar Säve-Söderberg. To judge by Jarvik's later reputation, he was thorough and brilliantly analytical, but he lacked flair or intuition. He tended to be dogmatic [J96] and somewhat unimaginative, at least by comparison to Säve-Söderberg. Säve-Söderberg seems to have been a golden boy -- charming, articulate, creative, and blessed with legendary energy [Jk96]. Thus Stensiö sent Säve-Söderberg to Greenland in 1930 and 1931 [LG04], rather than Jarvik. Consequently, it was S äve-Söderberg who found the first good tetrapod material from the region and published quickly, identifying two genera Ichthyostega and Ichthyostegopsis) and several species [LG04] [B05]. Jarvik was finally allowed to accompany Säve-Söderberg in 1932 or 1933. He found limited material of a new genus, which he later named Acanthostega [LG04] [B05].
 Säve-Söderberg continued collecting for several more years, but apparently put off monographic treatment until after his thesis was completed. As might be expected, Säve-Söderberg finished quickly and brilliantly, and he was immediately offered a professorship in 1937 [Jk96]. Unfortunately, that same year, Säve-Söderberg was struck by a mysterious wasting lung disease. Between his illness and the problems created by World War II, nothing was done about Ichthyostega. Eventually, Jarvik was ready to defend his own thesis in 1942 (he was 35 that year). Although Jarvik would continue to work in Stensiö's orbit for most of his life, Jarvik remained close to Säve-Söderberg, who was on his thesis Committee. Jarvik's thesis defense, however, was Säve-Söderberg's last public appearance. He died in 1948, and bequeathed the task of describing Ichthyostega to Jarvik [B05] [Jk96].
Säve-Söderberg continued collecting for several more years, but apparently put off monographic treatment until after his thesis was completed. As might be expected, Säve-Söderberg finished quickly and brilliantly, and he was immediately offered a professorship in 1937 [Jk96]. Unfortunately, that same year, Säve-Söderberg was struck by a mysterious wasting lung disease. Between his illness and the problems created by World War II, nothing was done about Ichthyostega. Eventually, Jarvik was ready to defend his own thesis in 1942 (he was 35 that year). Although Jarvik would continue to work in Stensiö's orbit for most of his life, Jarvik remained close to Säve-Söderberg, who was on his thesis Committee. Jarvik's thesis defense, however, was Säve-Söderberg's last public appearance. He died in 1948, and bequeathed the task of describing Ichthyostega to Jarvik [B05] [Jk96].
Thus, at the age of 41, after Jarvik was well-launched on his exhaustive studies of Eusthenopteron and other osteolepiforms, he was suddenly burdened with the responsibility for writing a definitive monograph on the most famous early tetrapod in the world, a creature wildly different from anything that had evolved before it -- and almost as different from the tetrapods which came after it. He was expected to do all this with Stensiö looking over his shoulder. Jarvik also knew (we may suppose) that (a) he was definitely not Stensiö's first choice, and (b) that he was supposed to fill the shoes of Säve-Söderberg, who had become the tragic hero of Swedish paleontology. Despite all this, Jarvik tried. He published a well-regarded paper on the post-cranial skeleton in 1952, and went on to lead at least one more collecting trip to Greenland. He also published a study of the limbs and limb girdles in 1980 [LG04] [B05]. However, all things considered, it isn't too surprising that he was less than eager to discharge this unwanted obligation. Unlike Wiglaf, Jarvik lacked the protection of Beowulf's shield -- nor did Wiglaf have the acerbic Stensiö [J96] offering helpful suggestions [2].
As a result, Jarvik was left to face the dragon alone, when he was in his eighties and without a Wiglaf of his own. One might argue that he ought to have let Clack and her students come in under his shield. Yet, from Jarvik's point of view, why would he want to pass the curse down to another generation? It was his wirgþu (curse, doom, punishment). But the saga-like irony of this course of action is that the curse has passed to one more generation. Ahlberg, Clack and co-workers are even now attempting the task of re-describing Ichthyostega because the current situation is untenable. Early tetrapod paleontology cannot continue with this key genus described with incomprehensible terminology, with many erroneous details, and with some parts never completed, almost 80 years after its discovery. Then, will the wirgþu continue? Who knows? But we hope Ahlberg et al. have fireproof shields this time, bring all their Wiglafs along -- and keep that wællseaxe handy. ATW050808.
[1] The phrase Forwrát ... wyrm on middan. Is usually rendered as "cut the dragon in two." For a variety of poetic, logical, cultural, and linguistic reasons, I'm convinced this is simply wrong.
[2] Our take on Stensiö's role is influenced by timing and the fact that Stensio was best known for his descriptive and theoretical work on the vertebrate skull, particularly the braincase [J96]. Although Jarvik attacked the axial skeleton of Ichthyostega with dispatch, he delayed his paper on limb girdles (recall that the pectoral girdle is part of the skull in fishes) until 1980, when Stensio was finally phasing out of active Museum work. Jarvik did not begin work on the skull proper until after Stensio's death in 1984, and never did much with the neurocranium.
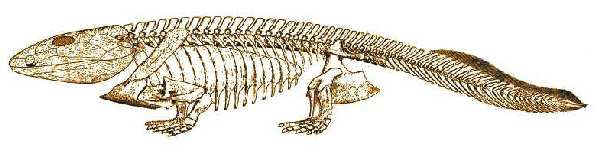 Ichthyostega: Säve-Söderberg, 1932. I. stensioei (stensiöi) Säve-Söderberg, 1932; I. eigili Säve-Söderberg, 1932; I. watsoni Säve-Söderberg, 1932 [B05].
Ichthyostega: Säve-Söderberg, 1932. I. stensioei (stensiöi) Säve-Söderberg, 1932; I. eigili Säve-Söderberg, 1932; I. watsoni Säve-Söderberg, 1932 [B05].
Late Devonian (Famennian) of East Greenland, Aina Dal & Britta Dal Fms. Possibly also Famennian of Scotland [A98] and France [Cle+04].
Phylogeny: Tetrapoda :: (Sinostega + (Densignathus + (Hynerpeton + (Tulerpeton + (Crassigyrinus + (Colosteidae + (Spathicephalus + (Baphetidae + Tetrapoda*)))))))) + *.
Characters: a robust Famennian tetrapod 1+ m long.
dermal skull: snout large & rounded; skull roof flat; unpaired median rostral [B05]; paired lateral rostrals present [R+03]; robust maxilla with anteriorly high portion forming triangle pointing towards jugal- lacrimal suture [B05]; nares between anterior tectal and lateral rostral; septomaxilla absent ( or = anterior tectal) [R+03]; prefrontal & jugal exclude lacrimal from orbit [R+03]; supratemporal pentagonal; intertemporal absent [Cl02a]; unpaired median postparietal [B05]; tabulars with posteroventral notch and deep ventrally projecting flanges attaching to the braincase; tabular suture with squamosal deeply interdigitated [R+03]; supratemporal & postparietal with large flanges extending downwards to receive braincase [Cl02a]; squamosal located relatively posteriorly (relative to parietal mid-length) [R+03];.
branchial: preopercular present [R+03]; deeply grooved gill bars for functional gills [Cl+03].
occiput: posttemporal fenestra absent [R+03] [2]; basioccipital was long and notochordal [R+03], with no occipital condyle;
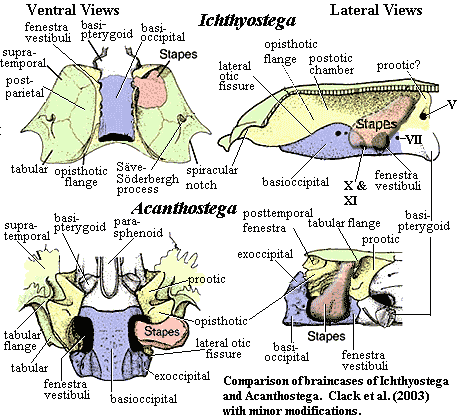 braincase: [1] notochord penetrates well into skull, as in Acanthostega [J+03]; otoccipital forming narrow crest suturing to median postparietal [Cl+03]; rounded sac-like structures flanking basioccipital; braincase long and narrow [$B05]; posterior stem of otic region very long and narrow [Cl+03]; opisthotics with large posterior "flanges suturing to down-turned extensions of the skull table margin" [$Cl+03]; prootics with strong transverse buttresses [Cl+03]; "On either side of the otoccipital braincase block is a large chamber, defined by solid walls formed of several different but closely sutured bones (proötic, opisthotic, epipterygoid and skull table ... ). The chamber extends above the otic capsules to the midline crest. ... Projecting into the chamber from the undersurface of the skull table is a finger-like scrolled structure named 'Säve-Söderberg's process'" [$Cl+03]; otic chamber homologous to middle ear [Cl+03]; stapes very "thin and leaflike, almost circular in outline and anterodorsally curved" [$B05]; sacculus chambers very small and anterior [Cl+03]; stapes with "'shaft' [] a very thin, almost circular, anterodorsally curved lamina of bone, projecting dorsolaterally into the otic chamber." [$Cl+03]; stapes with two heads, in contact but unfused, separated by large stapedial foramen [$Cl+03]; ventral cranial fissure not sutured [R+03]; basisphenoid does not suture to prootic or basioccipital [Cl98a]; basal articulation immobile [LCo95].
braincase: [1] notochord penetrates well into skull, as in Acanthostega [J+03]; otoccipital forming narrow crest suturing to median postparietal [Cl+03]; rounded sac-like structures flanking basioccipital; braincase long and narrow [$B05]; posterior stem of otic region very long and narrow [Cl+03]; opisthotics with large posterior "flanges suturing to down-turned extensions of the skull table margin" [$Cl+03]; prootics with strong transverse buttresses [Cl+03]; "On either side of the otoccipital braincase block is a large chamber, defined by solid walls formed of several different but closely sutured bones (proötic, opisthotic, epipterygoid and skull table ... ). The chamber extends above the otic capsules to the midline crest. ... Projecting into the chamber from the undersurface of the skull table is a finger-like scrolled structure named 'Säve-Söderberg's process'" [$Cl+03]; otic chamber homologous to middle ear [Cl+03]; stapes very "thin and leaflike, almost circular in outline and anterodorsally curved" [$B05]; sacculus chambers very small and anterior [Cl+03]; stapes with "'shaft' [] a very thin, almost circular, anterodorsally curved lamina of bone, projecting dorsolaterally into the otic chamber." [$Cl+03]; stapes with two heads, in contact but unfused, separated by large stapedial foramen [$Cl+03]; ventral cranial fissure not sutured [R+03]; basisphenoid does not suture to prootic or basioccipital [Cl98a]; basal articulation immobile [LCo95].
palate: palate closed; maxilla excludes premaxilla from choana [B05]; maxilla and premaxilla form shelf lingual to tooth row [R+03]; vomer sutures to maxilla anterior to choana [R+03]; vomer without anterior crest [R+03]; mesial margin of pterygoid, quadrate ramus, strongly anteriorly concave, forming large space lateral to braincase [$B05].
jaw: lower jaw robust with long dentary reaching articular [B05]; dentary loosely attached to surangular & angular, but firmly sutured to postsplenial & splenial [ACl98]; unique & complex symphysis formed of Meckelian bone [$ACl98]; parasymphysial plate present [R+03]; suture between prearticular and splenial deeply interdigitated [B05]; parasymphysial foramen on suture line with splenial [ACl98]; lateral parasymphysial foramen absent [ACl98]; coronoids large & wide [B05]; circular Meckelian foramina piercing wide strip of Meckelian bone [B05]; prearticular long & without shagreen; contact between prearticular angular + surangular not sutured [ACl98].
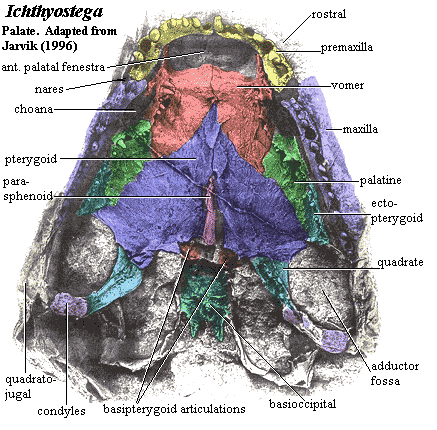 dentition:
~30 uniform dentary teeth [ACl98]; dentary teeth significantly larger than coronoid teeth, with very large symphysial fangs on dentary [B05]; dentary teeth widely separated [Cle+04]; no accessory dentary tooth row [ACl98]; parasymphysial fangs [R+03]; only one other tooth on parasymphysial plate [ACl98]; substantial gap between parasymphysial plate teeth and coronoid teeth [ACl98]; all coronoids with single marginal tooth row & without fang pairs, accessory tooth row, or denticle field [ACl98] [B05]; ~27 much larger teeth in upper jaw; maxillary tooth count 17-22 [B05]; maxillary teeth strongly curved, with posterior crest [B05]; vomerine, palatine & ectopterygoid fangs absent [R+03] [3]; weak posterior denticle fields on pterygoid [B05]; denticle fields generally absent from both upper & lower jaws [$ACl98].
dentition:
~30 uniform dentary teeth [ACl98]; dentary teeth significantly larger than coronoid teeth, with very large symphysial fangs on dentary [B05]; dentary teeth widely separated [Cle+04]; no accessory dentary tooth row [ACl98]; parasymphysial fangs [R+03]; only one other tooth on parasymphysial plate [ACl98]; substantial gap between parasymphysial plate teeth and coronoid teeth [ACl98]; all coronoids with single marginal tooth row & without fang pairs, accessory tooth row, or denticle field [ACl98] [B05]; ~27 much larger teeth in upper jaw; maxillary tooth count 17-22 [B05]; maxillary teeth strongly curved, with posterior crest [B05]; vomerine, palatine & ectopterygoid fangs absent [R+03] [3]; weak posterior denticle fields on pterygoid [B05]; denticle fields generally absent from both upper & lower jaws [$ACl98].
axial skeleton: vertebrae notochordal, but neural arches bore zygapophyses; neural spines squared off dorsally [Co96]; ribs long, blade-like, expanded & overlapping [B05]; cervical ribs distally expanded [R+03]; sacral rib present [Ca+05]; caudal fin with supraneurals & lepidotrichia (fewer & shorter than Acanthostega) [Co96] [Cl02a].
appendicular: cleithrum clavicle fused; anocleithrum absent [LCo95]; postbranchial lamina present [LCo95] [R+03]; clavicles do not meet anteriorly [R+03]; "posterior margin of interclavicle drawn out into parasternal process" [R+03]; large supraglenoid & supracoracoid foramina; moderately sized supraglenoid foramen within the triangular area [LCo95]; scapulocoracoid extends ventral to glenoid [R+03]; humerus L-shaped & flat; humerus without torsion [LCo95]; latissimus dorsi process weak or absent [LCo95]; humerus with prominent ectepicondylar buttress; ulnar & radial condyles well separated [LCo95]; ulnar condyle terminal; radial condyle ventral [A98] [R+03] (contra [Co+04], artifactual, citing unpublished recent work of others); forelimb with "permanently flexed elbow" [A98]; radius & ulna of equal length [Co+02]; ulna with conspicuous, forked olecranon [B05]; radius short & blunt [Co96]; manus unknown [Co+02]
pelvic: hindlimb flat and paddle- like [Co+02]; pelvic girdle large & unitary [Co+02]; pelvis with horizontal post-iliac process [A98]; & dorsally orientated iliac blade with unfinished facet for sacral rib attachment; extensive puboischiadic plate [Ca+05]; pubis sometimes separate from other elements [Co96]; acetabulum continuous with anterior edge of pelvis in most specimens [Co96]; supraacetabular buttress more prominent than postacetabular buttresses [Co96]; femur expanded proximally & distally [LCo95]; femur bearing large adductor blade [LCo95]; & deep intercondylar fossa; tibia large, quadrangular, with longitudinal ridge down medial surface [B05]; tibia & fibula both broad & flattened [LCo95] [Co+02]; tibia & fibula without well-defined shafts [Co96]; tibia significantly longer than fibula [Co96]; tibia with "well-developed cnemial crest, flanked anteriorly by a series of muscle scars, extends along the entire anterior edge of the lateral (extensor) surface" [LCo95]; fibula not waisted [R+03]; tarsus almost completely ossified [Ca+05]; large intermedium & fibulare present; fibulare large, articulating with two or more digits, as in Acanthostega [LCo95]; single? centrale [LCo95]; intermedium pentagonal [LCo95]; seven digits on pes [B05]; three small at leading edge & four stout digits more posteriorly [Co+02].
integument: ventral scales present [Ca+05].
Notes: [1] The braincase image requires explanation, since we have taken unusal liberties with the original in [Cl+03]. In addition to our usual complete relabelling, we have made the following modifications: (a) The Ichthyostega prootic is not identified in the original, and we've made our best stab at identifying it. (b) We've added a suture line between the tabular and supratemporal in the lateral view of Ichthyostega. c) In the ventral view of Acanthostega, we've changed some lines and coloring to clarify the shape and position of the tabular flange. (d) In the lateral view of Acanthostega we've added a complete stapes, including the presumed course of the unossified distal region, to conform to Clack's speculation that it connected with the spiracular notch. (e) In the same image, we've changed the shape of the tabular to reflect the tabular flange. Even with these changes, we still have concerns that the position and extent of the stapes is inconsistent between the two views of Ichthyostega. Finally, the anterior braincase of Ichthyostega is not shown, since it is largely unossified and unknown. [2] This part of the skull is so aberrant in Ichthyostega that it is not a simple matter to state whether a posttemporal fenestra exists or not. [3] !? see image.
Links: Ichthyostega; Devonian Times - about Ichthyostega; ICHTYOSTEGA AS A TRANSITIONAL FOSSIL; RE- Ichthyostega digits; ICHTHYOSTEGA; Ichthyostega stensioei; Palaeozoic puzzle; J17 Did labyrinthodonts evolve from lung fish or crossopterygians ... ; Biology 356.
References: Ahlberg (1998) [A98]; Ahlberg & Clack (1998) [ACl98]; Blom (2005) [B05]; Carroll et al. (2005) [Ca+05]; Clack (1998a) [Cl98a]; Clack 2002) [Cl02]; Clack (2002a) [Cl02a]; Clack et al. (2003) [Cl+03]; Clement et al. (2004) [Cle+04]; Coates (1996) [Co96]; Coates et al. (2002) [Co+02]; Coates et al. (2004) [Co+04]; Janvier (1996) [J96]; Jarvik 1996) [Jk96]; Johanson et al. (2003) [J+03]; Lebedev & Coates (1995) [LCo95]; Long & Gordon (2004) [LG04]; Ruta et al. (2003) [R+03]. ATW031004.
checked ATW040813