|
|
Bones: Dermal Bones |
| The Vertebrates |
Facial Series:
Premaxilla |
The Facial Series: Premaxilla
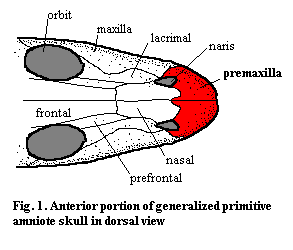 Anatomy: The premaxillae are paired dermal bones of the facial series, a series which also includes the maxillae and nasals. The morphology of the premaxilla is highly variable. It frequently takes the form of a 'C' in lateral view, with the top covering the tip of the rostrum, and the bottom forming the most anterior section of the upper jaw. It may be a major dermal component which plays a significant role in the margin of the nares (Fig. 1). Occasionally, as in hadrosaurs, the premaxilla is extensively elaborated in almost fanciful ways. See, for example, Hadrosauridae. In addition, the premaxillae may become invested externally with keratinous structures forming a beak or bill (rhamphotheca), as in the anomodont synapsids, birds, trilophosaurs and turtles Fig. 2). In these cases, the dorsal surface of the bone will often be roughened and show pits and channels. Where a rhamphotheca is absent, the premaxilla is usually a tooth-bearing bone. Most commonly, the premaxillae articulate with the maxillae and nasals, although borders with the frontals, prefrontals and other dermal bones are also possible.
Anatomy: The premaxillae are paired dermal bones of the facial series, a series which also includes the maxillae and nasals. The morphology of the premaxilla is highly variable. It frequently takes the form of a 'C' in lateral view, with the top covering the tip of the rostrum, and the bottom forming the most anterior section of the upper jaw. It may be a major dermal component which plays a significant role in the margin of the nares (Fig. 1). Occasionally, as in hadrosaurs, the premaxilla is extensively elaborated in almost fanciful ways. See, for example, Hadrosauridae. In addition, the premaxillae may become invested externally with keratinous structures forming a beak or bill (rhamphotheca), as in the anomodont synapsids, birds, trilophosaurs and turtles Fig. 2). In these cases, the dorsal surface of the bone will often be roughened and show pits and channels. Where a rhamphotheca is absent, the premaxilla is usually a tooth-bearing bone. Most commonly, the premaxillae articulate with the maxillae and nasals, although borders with the frontals, prefrontals and other dermal bones are also possible.
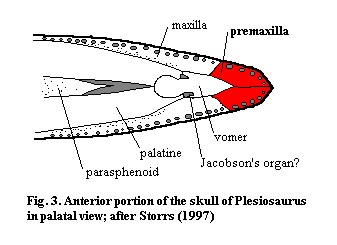
Ventrally, the premaxilla's tooth-bearing surface normally articulates with the vomers, dermal bones of the anterior palate, as well as the maxilla. It may also contact the palatines or their derivatives (dermal bones of the palate which are also closely associated with the maxilla) and even the parasphenoid. The premaxilla generally participates in the margin of the internal nares choana). As with the dorsal surface, the ventral premaxilla may also become a major element, as it is in birds. See, e.g., paleognathous bird palate. More typically, its ventral exposure is fairly limited (Fig. 3.). In a number of different groups, there is a gap in the tooth row, referred to as a diastema, corresponding to the articulation between the maxilla and premaxilla. This may be associated with a flexible premaxilla which is not completely sutured to the maxilla. See, for example, Ornithosuchus. Such an arrangement allows at least some independent movement of the tip of the rostrum. 
Functions: As largely discussed above, the premaxillae seem to be associated with the following functions:
1) Bearing the anterior-most teeth: for many carnivores, one might suppose that the front teeth would be of particular importance in capturing prey. While this may be true, the premaxillary teeth are not often specialized except in the therapsid line.
2) Supporting a beak. A wide variety of tetrapods have developed keratinous beaks for piercing or cutting food. Most are herbivores, but carnivorous birds and probably various closely related theropod dinosaurs also developed beaks.
3) Defining and supporting the chemosensory apparatus: The premaxilla normally defines a large portion of the naris and covers much of the nasal cavity.
 3)
Protection of the rostrum. For the simple reason that the premaxilla is usually the animal's most anterior bone, it is frequently the part that gets into trouble first. Quite likely this provided a powerful selective force favoring the formation of a fairly small, thick bone in this position, separate from the main body of the maxilla. In this way, an unexpected blow from the front is less likely to destroy the entire upper jaw or the structure of the face.
3)
Protection of the rostrum. For the simple reason that the premaxilla is usually the animal's most anterior bone, it is frequently the part that gets into trouble first. Quite likely this provided a powerful selective force favoring the formation of a fairly small, thick bone in this position, separate from the main body of the maxilla. In this way, an unexpected blow from the front is less likely to destroy the entire upper jaw or the structure of the face.
4) Specialized structures. In fish, the premaxilla is an important element in a number of different arrangements which allow protrusion of the mouth to suck in and envelop prey (see, infra). As noted above, the premaxilla may also become involved in the structure of facial ornament such as crests and the elaboration of the nasal passage in hadrosaurs.
Phylogeny: Every elongate organism has to have a front end. So, in a sense, analogs of the premaxilla go as far back as chordates with bone. However, the premaxilla, even considered as a functional unit of the skull and without regard for phylogeny, is a much more complicated object. As fully developed in advanced tetrapods, it has a three-fold structure. (1) Ventrally it is involved with prey capture and dentition. (2) Dorsally, it protects the rostrum, a portion of the body far more likely than most to suffer damage. (3) Internally, it is involved with respiration and chemosensation. In a very broad sense, the premaxilla seems to have acquired these functions in the order listed.
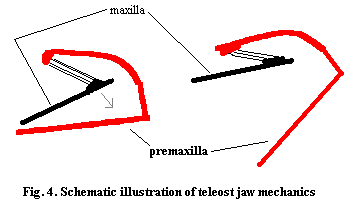 In Cheirolepis, one of the most basal animals with more or less "standard" bones, the premaxillae appear to have been primarily tooth-bearing bones. Externally, they also formed the broad base of a fairly tall rostrum dominated by the rostral bones and a series of small dermal bones with no clear correspondence to those of living species. In more derived actinopterygians, the orbit is much further back on the skull so that the rostrum is no longer vertical, but slopes gradually upward from the terminal mouth. As a result, the external exposures of the premaxillae are naturally stretched out and angled back from the mouth. During the same evolutionary sequence, the premaxillae generally became the principle tooth-bearing element of the upper jaw, while the maxilla developed into a lever, structured so as to protrude the premaxilla and drop the floor of the mouth for suction feeding. As this arrangement requires a mobile premaxilla, things get quite complicated. One might anticipate that the premaxillae would then lose their dorsal, external exposures. Instead, the "ascending processes" are retained, although modified, and become part of a lever used to swing the ventral, internal, portion of the premaxilla down and out. The mechanics of this type of arrangement are illustrated (rather inadequately, to be sure) in Figure 4.
In Cheirolepis, one of the most basal animals with more or less "standard" bones, the premaxillae appear to have been primarily tooth-bearing bones. Externally, they also formed the broad base of a fairly tall rostrum dominated by the rostral bones and a series of small dermal bones with no clear correspondence to those of living species. In more derived actinopterygians, the orbit is much further back on the skull so that the rostrum is no longer vertical, but slopes gradually upward from the terminal mouth. As a result, the external exposures of the premaxillae are naturally stretched out and angled back from the mouth. During the same evolutionary sequence, the premaxillae generally became the principle tooth-bearing element of the upper jaw, while the maxilla developed into a lever, structured so as to protrude the premaxilla and drop the floor of the mouth for suction feeding. As this arrangement requires a mobile premaxilla, things get quite complicated. One might anticipate that the premaxillae would then lose their dorsal, external exposures. Instead, the "ascending processes" are retained, although modified, and become part of a lever used to swing the ventral, internal, portion of the premaxilla down and out. The mechanics of this type of arrangement are illustrated (rather inadequately, to be sure) in Figure 4.
The sarcopterygian premaxilla appears to be homologous to the actinopterygian bone, although the case is not as clear as it might be. Some dipnomorphs, for example, develop a fused parietoethmoidal shield, which unites the parietals with the rostral bones in a single plate. Others break up the dermal skull in ways which bear no relation to the standard pattern. However, the most common arrangement of the premaxillae is quite similar to Cheirolepis. The significant difference is that the sarcopterygian orbit is so far forward that the nares are brought into close proximity with the premaxilla. In addition, the sarcopterygian brain is sharply divided anteroposteriorly, with the anterior ethmoid practically pushed against the internal surface of the premaxilla. In tetrapods, the premaxillae frequently participate in the margin of the internal nares, the choana. However, this appears to be a late development. The choanae originally opened at the intersection of the vomer, palatine and maxilla -- similar, and perhaps homologous to the vomeronasal or Jacobson's organ of the later snakes and plesiosaurs.
From that point forward, the premaxilla has been extraordinarily stable. In some groups with highly consolidated skulls, such as turtles, the premaxilla is small or disappears. In others, as noted above, it becomes prominent. Generally speaking, it seems to have taken over a larger role as a rostral bone, and lost ground in the palate. This trend is especially clear in archosaurs, but is reversed in birds. On the whole, however, there is remarkably little change in form or function, except perhaps in highly derived snakes. Even in mammals, the relative position and articulations of the premaxillae are quite similar to those in sarcopterygians, allowing for the sloppy tendency of mammals to fuse everything together.
This series of essays is far too new at this date (November, 2000) to be drawing conclusions, but the extraordinary stability of this bone calls for some explanation. Perhaps (and solely as a tentative hypothesis going forward) diversity of function generates phylogenetic stability. The premaxilla is not only integrated with a dermal skull "program." It is deeply involved in the prey capture and mastication programs, as well as the chemosensation system. Thus different developmental genetic pathways are so deeply entangled at this junction that it becomes very difficult for mere selection or genetic drift to cause really significant change in the morphology of this bone. ATW001103.
checked ATW050523