 G. Anatomical Correlates of the Inner Ear in Reptiles
G. Anatomical Correlates of the Inner Ear in Reptiles| Bones: The Ear | ||
| The Vertebrates | Ear Overview (2) |
| s ("Cladograms") |
||||||
| Vertebrates Home | Vertebrate | Vertebrate | Bones | Time |
| Ear Overview The Incus References |
What about the blue area, the scala tympani? One design problem for the perilymphatic duct is that it is very small, and entirely too efficient at capturing the energy generated at the fenestra ovalis. If the perilymphatic duct simply hit a dead end, any sufficiently loud sound would reflect back along the duct, causing untoward interference waves -- or the perilymphatic duct might simply explode. To prevent this, the duct has to dissipate excess sound energy which has reached the end of the lagena. Physics allows us two ways to do this, both of which are also used by university administrations to control the excess energies of noisy faculty committees. Either (a) the noisy vibrations (or faculty) are allowed to bounce around randomly in an enclosed space, until their excess energy is used up in mutual interference and frictional heat, or else (b) the vibrations are used coherently to perform some important-looking, but actually useless work, such as pushing on elastic membranes or piles of reporting forms, until they are exhausted.
The scala tympani uses both approaches, to different degrees in different lineages. Basal reptiles, even those without an elongate lagena, simply run the perilymph into a large sinus in the brain or in the metotic fissure between the otic capsules, where compression waves bounce around and dissipate like echoes in a cavern. Other organisms tend to combine this strategy with an extension of the perilymphatic duct which runs out through the metotic foramen and ends at an elastic membrane, the fenestra rotunda. Since the fenestra rotunda is, properly speaking, a specialized mammalian structure, the membrane is frequently referred to as the fenestra pseudorotunda in reptiles. This fenestra faces out into some effectively infinite pressure sink, such a Eustachian tube connected to the pharynx.
Mammals, as we have learned to expect, employ a bizarre and pointlessly complicated variation on the straightforward and logical reptilian plan. The mammalian lagena, called a cochlea, is spiral-shaped and drills into the surrounding bone like a corkscrew opening some debased Australian vintage stoppered with bone. The perilymphatic duct has, of course, no choice but to follow along. The problem is that, at the end of the spiral tunnel, the perilymphatic duct finds itself deep inside the temporal bone, with no place to go. Perhaps realizing its tactical error, the duct sheepishly turns around and stumbles back out the way it came, but on the other side of the cochlea, following the basilar membrane.
G. Anatomical Correlates of the Inner Ear in ReptilesSometimes we are lucky enough to find nice fossil impressions of these inner ear structures. Then again, if we were generally that lucky, we would be living on a yacht off some notorious Caribbean tax haven, surrounded by fawning sycophants and unimaginable luxuries, enjoying the myriad fruits of our multiple lottery winnings. Fate is rarely so generous. We have to make do with bones for the most part. Fortunately, there are some hard tissue correlates which can sometimes give us an idea of what was going on inside the inner ear.
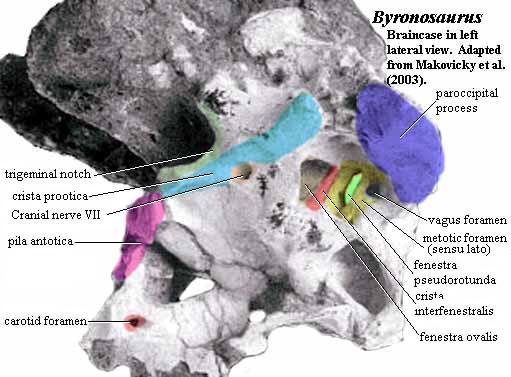
One such outward sign is the crista interfenestralis, shown in the braincase of the troodont, Byronosaurus. Makovicky et al. (2003). In the image, we have included some other landmarks which may be familiar from our discussion of the generalized amniote braincase. However, the point of the image is the crista interfenestralis which divides the inner ear into two parts. Other structures or the same structure with other names) may perform the same job. For example, in the crurotarsan Batrachotomus, shown below, the inner ear is divided by a ventral ramus of the opisthotic (dark blue). The crista interfenestralis and the ventral ramus of the opisthotic may, in fact, be the same thing.
However the inner ear is divided, the osteological result is the same. On the anterior side is the fenestra ovalis, where the columella (or stapes or hyomandibular, depending on the taxon and your tastes in nomenclature) makes contact with the inner ear. The fenestra ovalis frequently has a triangular shape, as in Byronosaurus. In organisms with an elongate lagena, we will observe a long lagenar recess, since the hearing mechanism for these animals requires a fixed basilar membrane and a carefully controlled interface with the perilymphatic duct. Sometimes, too, an additional bone (the crista vestibuli) walls off the lagena + perilymphatic duct from the rest of the vestibular apparatus. The functional significance of this wall is probably both structural stability and acoustic insulation. The presence of such a barrier bespeaks a lifestyle which requires both acute hearing and a high degree of pitch discrimination.
 On the posterior, scala tympani side of things is a whole complex area of foramina which we will refer to as the metotic foramen, sensu lato. This posterior region requires some energetic sorting out, and we will now set about that task.
On the posterior, scala tympani side of things is a whole complex area of foramina which we will refer to as the metotic foramen, sensu lato. This posterior region requires some energetic sorting out, and we will now set about that task.
Embryologically, the metotic fissure is the gap between the developing otic placodes and the parachordal bars which serve as the foundation for the entire head posterior to the sella turcica. All of the cranial nerves exiting the posterior part of the brain exit into this space, particularly, cranial nerves IX, X, XI & XII, as well as some of the important veins which drain the braincase. This is also the space in which the perilymphatic duct terminates, in more primitive vertebrates, with a dead end cistern used to absorb excess noise. In early development, there's lots of room here. Only the ventral part is closed off, by the parachordal mesoderm. So, what happens when, over the course of development (and evolution), the following happen:
(a) the anterior end is closed by the lateral commisure, the joining of the otic capsules to the brain;
(b) the dorsal part is closed off by various outgrowths of the capsules (the supraoccipital, epiotic, and membranous precursors) as well as the dermal skull bones;
(c) the posterior part is filled up and blocked off by the recruitment of embryonic proto-vertebrae to form the occipital bones; and
(d) everything grows and expands?
What happens is that the animal simply runs out of room. Everything gets squeezed into a little metotic foramen (the purple area in the figure of Batrachotomus). There's no room for a perilymphatic cistern, so excess pressure has to be relieved some other way -- a  fenestra rotunda or equivalent. Initially, at least in an evolutionary sense, everything tries to escape by this same door, the metotic foramen. Eventually, there's no room for the nerves or veins to exit either, so they find other ways out. Typically, they develop their own foramina between or around the embryonic neural arches which form the exoccipitals. The most prominent of these exits is the vagus foramen, homologous to the jugular foramen or "posterior lacerate foramen" of mammals. Quite often the hypoglossal (XIIth) nerve develops its own exit, the hypoglossal foramen.
fenestra rotunda or equivalent. Initially, at least in an evolutionary sense, everything tries to escape by this same door, the metotic foramen. Eventually, there's no room for the nerves or veins to exit either, so they find other ways out. Typically, they develop their own foramina between or around the embryonic neural arches which form the exoccipitals. The most prominent of these exits is the vagus foramen, homologous to the jugular foramen or "posterior lacerate foramen" of mammals. Quite often the hypoglossal (XIIth) nerve develops its own exit, the hypoglossal foramen.
When most of the nerves, veins and other riff-raff have been evicted, the remnants of the old metotic foramen can begin to relax. Since it no longer has to accommodate a group of distracting nerves and veins, this fenestra can now specialize to dissipate excess sound energy, usually by pushing a membrane into an external pressure sink, as discussed above. At this point, we may refer to the metotic foramen as the fenestra rotunda -- or, more precisely, as a fenestra pseudorotunda in reptiles.
The function of the inner ear is, then, to package an enormous amount of information from mechanoreceptors into a form usable by the brain or, in some cases, directly by motor neurons. It transduces various inputs into neural electrical signals. The function of the middle ear and of certain functionally analogous structures is also signal transduction. In this case, the signals are transduced from vibrations, usually airborne, to vibrations in the fluid medium of the perilymph in the inner ear. Fish, of course, don't have this problem. The sounds of concern to them are largely water-borne. However terrestrial animals must get vibrational information from the air to the mechanoreceptors in the fluid-filled inner ear.
The principle difficulty here is that water is different from air. The standard textbook explanation invokes density, the implication being that it takes relatively more energy to get a fluid vibrating, simply because the medium is denser. But, even more importantly, air and water are chemically different. The modes, characteristic frequencies, and speed of propagation of vibration differ between the two media. As anyone knows who has tried to hear while diving, most airborne sound is reflected from the surface of a fluid (or dissipated as random heat). This is the difference in acoustic impedance which is actually what is invoked in the phrase "impedance-matching ear." The design problem for a terrestrial organism is to evolve an ear that transduces vibration from air to fluid over the considerable barrier posed by the difference in impedance.
Figure 4 shows, in very schematic form, how the trick is done. The tympanic membrane receives the airborne vibration. Unlike a fluid surface, it is locally more rigid. Thus it responds as a unit to average air pressure changes over a considerable distance. Much acoustic energy is reflected away still, but the microscopic oscillation of individual air molecules has been traded for a macroscopic and highly coordinated mechanical movement of the membrane surface. Thus, despite a serious loss of total signal energy, the signal to noise ratio is very favorable and the signal energy has been captured in a form available to do mechanical work.
These macroscopic movements of the tympanum are transmitted through the columella and extracolumella in most sensible tetrapods, or through a Rube Goldberg machine of three weirdly shaped bones in mammals. In either case, the system is in part designed to achieve a high ratio between the area of the tympanum and the area of the fenestra ovalis. This achieves an impressive amplification of the signal. The columella is attached to the fenestra ovalis by flexible ligaments which allow the movement of the bone to transmit a corresponding vibration to the fluid surrounding the vestibular apparatus.
[Speculation: presumably, the system has also evolved to transmit information about high frequency overtones. A two-dimensional resonator, such as the tympanum, does not simply vibrate in and out as a whole. Overtones are carried as a complex pattern of vibration, with the surface partitioned into different areas moving in different directions. The length of the columella, as well as the projections from its contact with the tympanum, appear to allow it to transmit information about such overtones as lateral forces.]
Since water-based fluids are incompressible for all practical purposes, the inner ear also has a second membrane-covered window, the fenestra rotunda "round window") which flexes in the opposite sense from the base plate of the columella (or the stapes in mammals). Pressure changes in the middle ear itself are accommodated through the Eustachian tube which communicates with the throat, as discussed in a previous section.
In fish (which have no middle ear) and amphibians, some taxa have developed additional devices for transmitting sound to the inner ear. Some fish, for example, have processes from the swim bladder to the inner ear to transmit airborne vibrations. Others have developed a series of small bones, the Weberian ossicles, which perform the same function. In amphibians, the columella shares the oval window with a second bone, the operculum, which communicates by way of an opercular muscle with the pectoral girdle. Apparently, this is the primary route of transmission of the "seismic sense" in basal tetrapods.
Our tour of the vertebrate ear ends with the outer ear, which seems to be fairly uninteresting from a phylogenetic perspective. In mammals, the ear is typically ornamented with a cartilaginous pinna (what we colloquially refer to as the "ear"). This optimizes the external channel that sound follows to reach the tympanum, which is referred to as the external auditory meatus, so that sound is focused on the tympanic membrane and the directionality of the signal is optimized.
ATW001113, last revised ATW031223.
Hearing in Stegocephalians; Morten Buhl Jørgensen's homepage; What is acoustic impedance?; Surgical Technique of Jean-Bernard CAUSSE, M.D. (1/4); Anatomy and Physiology; Music for the sacculus: A blast from the past; Springer LINK: Pflügers Archiv - Abstract Volume 435 Issue 1 (1997) pp 82-90; Grand_Rounds ... Vestibular_Physiology_1992 (very nice, but unillustrated, tour of the human vestibular apparatus); Histology and Microscopic Anatomy of the Ear.
1. Incus
[1] Not always tiny.Some coelacanths have very large otoloths for reasons which are not only obscure, but hard to understand on functional grounds.
[2] All in the Early Triassic, it would appear -- which is downright odd. Was there something in the nature of the End-Permian extinction which selected for terrestrial vertebrates with pitch discrimination, or at least high-frequency hearing?
last modified ATW060131
checked ATW050819