
Panderichthys, from the late Devonian of Latvia. Artwork by Nobu Tamura, 2007. Wikipedia, GNU Free Documentation/Creative Commons Attribution license.
| Sarcopterygii | ||
| The Vertebrates | Panderichthys, etc. |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Teleostomi
├─Neopterygii
└─Sarcopterygii
├─┬─Onychodontiformes
│ └─Actinistia
└─Rhipidistia
├─Dipnomorpha
└─Tetrapodomorpha
├─Rhizodontiformes
└─Osteolepiformes
├─Tristichopteridae
└─Elpistostegalia
├─Panderichthys
└─┬─Elpistostege
└─┬─Livoniana
└─┬─Elginerpetonidae
└─┬─┬─Metaxygnathus
│ └─Ventastega
└─Tetrapoda
|
Contents
Overview |
Panderichthys, from the late Devonian of Latvia. Artwork by Nobu Tamura, 2007. Wikipedia, GNU Free Documentation/Creative Commons Attribution license. |
We've identified two fundamental adaptive trends in rhipidistian evolution, and really only two. To be fair, we use "trend" here in a tightly constrained sense. An adaptive trend must exhibit sustained, directional development independently in multiple lineages. Our first trend, the development of a strong anteroposterior arch from snout to occiput, is as well evidenced as these things are ever likely to be. The second, a distinctive, tetrapod-like locomotor style, is fuzzier, but explains much that is otherwise hard to understand. The two developments are probably somewhat related. If the forelimb takes on a significant locomotor role, it is at least somewhat less available for steering and attitude control. A large, flat skull with some mobility and a strong central brace can help to make up for the lack. This scenario would provide a straightforward rationale, for example, for the evolution of the peculiar "neck" of Mandageria and the "eyebrows" of the Elpistostegalia. Both can easily be related to directional control in swimming. As we have noted, the development of hind limbs is also an expected and natural development helping to free up the forelimbs for their original function.
We were originally dubious of the theme stressed by Clack, Ahlberg, Johanson and others. These workers emphasize that most of the major tetrapod innovations are adaptations for a particular aquatic lifestyle and not adaptations to terrestriality. The concept is counterintuitive. Nevertheless, our review of rhipidistian anatomy suggests that this may be exactly what happened. With that said, we still leave a deeper question unanswered. These are adaptive trends. They do not seem to represent niche specializations, precisely because the overall pattern is independent of lineage. What were the rhipidistians adapting to?
One set of possibilities revolves around the idea that, even though we observe aquatic adaptations, this particular set of adaptations was convergently acquired because they were compatible with occasional exposure to a terrestrial environment. There are two serious anatomical problems with this line of speculation: fingers and brains. The evolution of digits lagged almost every other tetrapod innovation. Likewise, the internal obliteration of the intracranial joint is only observed in tetrapods. Ahlberg et al. (1996).
We believe that these are serious criticisms, but not sufficient to outweigh the obvious logical advantages of invoking dry land to explain adaptations to dry land. As to fingers, if we were to find a fish with fingers, we would probably revise our ideas about Tetrapoda, rather than our ideas about phylogeny. That is, only tetrapods have digits because the label "tetrapod" has historically been set as a marker for things with digits. Perhaps more importantly, there may well be anatomical reasons why digits were late-appearing adaptations. Making any significant use of digits requires an elaborate set of muscles, nerves and receptors. Not only is this a complex series of adaptations, but it may not be compatible with the basic propulsive design of these limbs. As the actinopterygians "learned," lightweight fin rays are so much more efficient than bone for distal limb elements, that it makes no sense to place bone in a limb if it can be avoided. Thus, the late development of digits may be reasonably explained.
But if this is correct, what about rhizodonts? They have digit analogues, and (in spite of our earlier comment) are not really good candidates for terrestriality. Possibly, it was simply the great size of these fishes that made lepidotrichia impractical. Another possibility is that cartilaginous plates on the distal limb were relatively common but ossified only in very large fishes like rhizodonts. At this point, too little is known of the rhizodonts to make too much of this character.
The persistence of the divided braincase is likewise not too troubling. The dermal joint was lost in the Elpistostegalia. In Mandageria, the dermal joint was plainly present, but some of the area ventral to the braincase, an area which had housed the muscles controlling the joint in basal taxa, was recycled to provide the "neck." Thus, in at least these two lineages, the divided braincase already had reduced functional significance. However, there are at least no obvious disadvantages to the divided braincase until the loss of the opercular region and cranial shoulder girdle reduced the lateral and posterior support for the otoccipital region. At that point -- the level of Acanthostega -- it becomes reasonable to suppose that this antique division of the brain first became a liability.
We return now to this famous phrase of Romer's. We do so fully conscious that this is just story-telling, as we said at the start. The purpose is to focus on the environmental aspects of tetrapod evolution which should not be forgotten, even if we can never be sure exactly what the relevant environment was. Human beings being what they are, we all tell stories to ourselves. We believe that it furthers the goals of science to tell those stories explicitly, rather than pretend that we have no preconceptions.
In 1980, Phillipe Janvier (1980) published a brief paper with preliminary observations on some osteolepiform shoulder girdles found at various places in Iran and eastern Turkey. The paper has managed to be both obscure and classic. It is obscure in that the specimens, so far as we know, are still undescribed. The Devonian exposures in the region are not well-studied, and the taxonomic position of these fishes can't be determined with any certainty. It is classic, and frequently cited, because it is vintage Janvier -- full of interesting ideas and delight at poking around for new fossils in odd corners of the world. One of the observations he makes is that the world saw a marked increase in the area covered by tidal flats in the Middle and Late Devonian. Certainly sea levels were high and the geological record suggests that weathering rates had increased. Perhaps just as importantly, marginal marine plants were now common, fixing and stabilizing sediment along the shorelines. Some casual research in another direction suggests that tidal range was also markedly (~25%) higher than today, simply by virtue of long-term changes in the position of the moon.
Building on these concepts, one wonders whether there might be something to the old saw of Romer's after all. The "periodic droughts" might have occurred every twelve hours or so at low tide, or twice a month at neap tides, or during some dry season annually. It isn't necessary to invoke rare or cataclysmic events. Animals living on or around tidal flats, estuaries, coastal bars, and barrier islands will always experience periodic low water. The larger members of these communities usually retreat with the tide. Yet, sometimes, they can't or don't. This was a time without birds or mammals, and the trophic rewards might be high for an animal that could take up the easy gleaning of stranded fishes or prey caught in pools by the low tide.
As for periodic droughts, they happened, too. Look at Canowindra. There are tetrapod tracks in Late Devonian Australia, but there are no elpistostegalians or tetrapods at Canowindra. Perhaps they didn't live there. But, then again, maybe they just walked away. ATW031109.
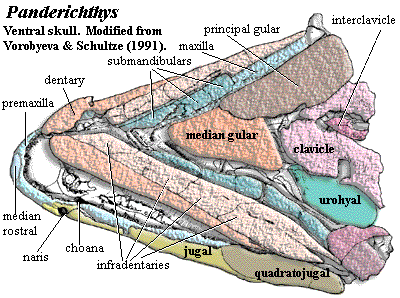 Panderichthys: Gross, 1941. P. rhombolepis Gross, 1930; P. stolbovi Vorobyeva, 1960.
Panderichthys: Gross, 1941. P. rhombolepis Gross, 1930; P. stolbovi Vorobyeva, 1960.
Late Devonian (Frasnian) of Europe (Latvia) & Russia [CA96]
Elpistostegalia:: Elpistostege + *.
90-130 cm long [V80]; Cranial: head long, ~25% body length [V80]; head dorsoventrally flattened, triangular, narrow at the snout and wide posteriorly; mouth subterminal [V80] [SA85]; well-developed rostrum anterior to nares [V80]; unpaired median rostral [VS91]; rostrum ventrally curved [VS91]; maxilla narrow [VS91]; nares marginal [VS91]; nares enclosed by anterior tectal dorsally & a narrow lateral rostral ventrally [VS91]; very thin posterior process of premaxilla separates nares from jaw line [VS91]; nares with well-developed dermintermedial process [V80]; dermintermedial process divides nares horizontally [VS91] [1]; nares larger than in Eusthenopteron and closer to jaw [C02]; 3 pairs of nasals [SA85]; paired frontals present [V80] [VS91]; unpaired median postrostral may be present [VS91]; postfrontal ridge ("eyebrows") medial to orbits [SA85] [VS91]; parietal meets frontal between orbits [SA85]; narrow anterior portion of parietals between orbits [VS91]; parietal with small pineal foramen posterior to orbits [VS91]; parietal-postparietal suture serrated and akinetic [V80] [4]; postparietals may be fused to median extrascapular [VS91]; tabular & supratemporal lateral to postparietal [VS91]; extrascapulars small [VS91]; lateral extrascapulars overlap median extrascapular [VS91]; 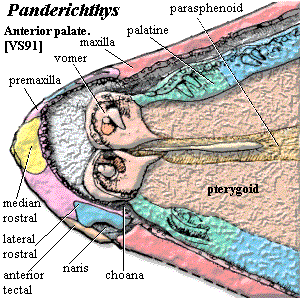 tabular separated from cheek by long spiracular slit [VS91]; spiracular slit reaches posterolateral corner of supratemporal [VS91]; postfrontal sutures to intertemporal posteriorly [VS91]; intertemporal present [SA85]; long lacrimal joins anterior tectal and orbit, lateral to prefrontal [VS91]; orbits located high on skull and close together [VS91]; orbits posteriorly placed, transversely elongate [V80]; prefrontal & postfrontal form anterior & dorsal margins of orbit [VS91]; jugal reaches orbit [SA85]; jugal & postorbital long, with jugal forming most of postorbital cheek [VS91]; cheek consolidated with skull roof, without lateral kinesis [V80] [VS91]; squamosal large [VS91]; squamosal meets maxilla, separating jugal from quadratojugal in some specimens, but jugal meets quadratojugal one side of one specimen [VS91] [C02]; opercular series much shorter anteroposteriorly than in Eusthenopteron [C02]; quadrate double-headed [VS91]; Braincase & PQ: palatoquadrate without lateral ethmoid articulation [V80]; palatoquadrate has synchondral articulation with ethmoid [VS91]; hyomandibular flat, elongate, straight, lying in depression on dorsomedial surface of palatoquadrate [VS91]; ventral hyomandibular articulation displaced posterior to dorsal articulation [J+03]; vestibular fontanelle small and anteriorly placed [A+96] [J+03]; intracranial joint present in braincase but lost from the external skull [A+96]; intracranial joint not significantly different from basal rhipidistians [A+96]; nasal capsules small & well-separated [V80]; posterior wall of nasal capsule unossified [VS91]; Palate: large anterior palatal pit for dentary fangs [VS91]; premaxillary palatal plate absent [VS91]; vomers with fanged plates, meeting medially [VS91]; choana present, with distinct passage to nasal capsules [VS91]; choana elongate, with wide anterior portion between vomer & premaxilla, elongated posterior between palatine & maxilla [VS91]; parasphenoid narrow, flanked by short vomer posterior processes [VS91] [JA97]; parasphenoid dorsal body connected to tooth-bearing plate by narrow septum [VS91]; interpterygoid vacuity absent, with pterygoid covering this region [VS91];
tabular separated from cheek by long spiracular slit [VS91]; spiracular slit reaches posterolateral corner of supratemporal [VS91]; postfrontal sutures to intertemporal posteriorly [VS91]; intertemporal present [SA85]; long lacrimal joins anterior tectal and orbit, lateral to prefrontal [VS91]; orbits located high on skull and close together [VS91]; orbits posteriorly placed, transversely elongate [V80]; prefrontal & postfrontal form anterior & dorsal margins of orbit [VS91]; jugal reaches orbit [SA85]; jugal & postorbital long, with jugal forming most of postorbital cheek [VS91]; cheek consolidated with skull roof, without lateral kinesis [V80] [VS91]; squamosal large [VS91]; squamosal meets maxilla, separating jugal from quadratojugal in some specimens, but jugal meets quadratojugal one side of one specimen [VS91] [C02]; opercular series much shorter anteroposteriorly than in Eusthenopteron [C02]; quadrate double-headed [VS91]; Braincase & PQ: palatoquadrate without lateral ethmoid articulation [V80]; palatoquadrate has synchondral articulation with ethmoid [VS91]; hyomandibular flat, elongate, straight, lying in depression on dorsomedial surface of palatoquadrate [VS91]; ventral hyomandibular articulation displaced posterior to dorsal articulation [J+03]; vestibular fontanelle small and anteriorly placed [A+96] [J+03]; intracranial joint present in braincase but lost from the external skull [A+96]; intracranial joint not significantly different from basal rhipidistians [A+96]; nasal capsules small & well-separated [V80]; posterior wall of nasal capsule unossified [VS91]; Palate: large anterior palatal pit for dentary fangs [VS91]; premaxillary palatal plate absent [VS91]; vomers with fanged plates, meeting medially [VS91]; choana present, with distinct passage to nasal capsules [VS91]; choana elongate, with wide anterior portion between vomer & premaxilla, elongated posterior between palatine & maxilla [VS91]; parasphenoid narrow, flanked by short vomer posterior processes [VS91] [JA97]; parasphenoid dorsal body connected to tooth-bearing plate by narrow septum [VS91]; interpterygoid vacuity absent, with pterygoid covering this region [VS91]; 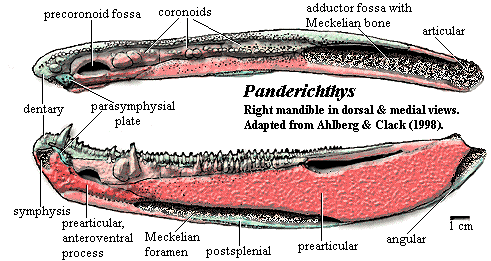 Jaw: jaw generally like basal osteolepiforms [AC98]; outer face of lower jaw gently curving & flattish [AC98]; posterior jaw formed by surangular without surangular crest [AC98]; lower jaw with long dentary and 4 infradentaries [VS91] [2]; lingually, jaw with prearticular and 3 coronoids [VS91]; prearticular extends anteriorly to symphysis, overlain by parasymphysial plate anteriorly [VS91]; prearticular forms medial edge of adductor fossa [AC98] [6]; prearticular with anteroventral process forming medial/ventral edge of precoronoid fossa [AC98]; ventral margins of prearticular & infradentaries held together by strip of Meckelian bone [AC98]; prearticular anteroventral process partially separated (might be a separate ossification), and underlies anterior of prearticular [AC98]; parasymphysial plate small, not contacting coronoid [JA97]; Meckelian completely ossified, exposed at symphysis and in all fossae (see image) [AC98] [7]; articular continuous with Meckelian [AC98]; parasymphysial plate with irregularly spaced small teeth [AC98]; dentary with homodont teeth, some specimens with additional smaller outer row [AC98]; dentary with fang pair posteromedial to symphysis [AC98]; coronoids with pits accommodating upper fangs [VS91]; fangs on coronoids 1-2, variably on 3 [AC98]; teeth polyplocodont, with minimal bone of attachment [VS91]; large median gular and large principal lateral) gulars [V80] [SA85]; 7 submandibulars [VS91] [3]; Axial: body narrow & elongate [V80]; vertebral column ossified throughout; centra ring-like (except for ventral slit) near pectorals and caudal fin [V80]; centra made up of large intercentra fused with small pleurocentra [V80] (contra [VS91]: pleurocentra absent); neural arches sutured to dorsal border of intercentra, do not meet or fuse, neural spines paired [VS91]; short, broad dorsal ribs attached (continuously) to arch and intercentrum [V80] [VS91] [5]; enlarged ribs which project ventrally (?!); fin basal scutes absent [V80]; dorsal and anal fins absent or confluent with caudal [V80]; reduced, diphyceral tail [V80]; caudal fin with surrounding finfold [VS91]; Appendicular: dermal shoulder girdle attached to skull through usual series of cleithrum, anocleithrum, supracleithrum & posttemporal [VS91]; clavicles broad & broadly in contact, with small posteromedial interclavicle [VS91]; clethrum with groove to receive ascending process of clavicle [C96]; scapulocoracoid massive, with single, broad attachment to cleithrum [V80] [VS91] [C+02]; probably no scapular blade [C+02]; scapula thickened at attachment to cleithrum [C96]; coracoid plate large, with very large supracoracoid foramen [VS91] [C+02]; glenoid fossa as shallow groove (exact shape unknown) [VS91]; pectoral fins positioned far ventrally [V80] [VS91]; pectorals with 3- mesomeres, with each set specialized, not clearly segmented [S95] [C+02]; pectoral fin radials absent [S95]; humerus elongate (by sarcopterygian standards) [V80]; humerus dorsoventrally flattened, with distinct extensor & flexor surfaces [C96] [C+02] [J+03]; humerus with well-developed deltoid crest [VS91]; entepicondylar postaxial) process absent [VS91] (contra [C+02]: present and dorsoventrally flat); radial condyle ventral [C+02] (contra [S+04]: distally facing) ; radius as long as humerus, with well-developed ventral ridge [VS91]; ulna 50-75% as long as humerus [VS91]; distal forelimb with intermedium and plate of fused radial bones (ulnar plate), not digits [V80]; no distal radials on plate ("ulnare") [C+02]; forelimbs with numerous lepidotrichia; lepidotrichia branched [VS91] pelvics small, set far posteriorly, near caudal fin [V80] [VS91]; Integument: scales small & without cosmine [VS91]; ornamentation on scales tuberculate & ridged [VS91].
Jaw: jaw generally like basal osteolepiforms [AC98]; outer face of lower jaw gently curving & flattish [AC98]; posterior jaw formed by surangular without surangular crest [AC98]; lower jaw with long dentary and 4 infradentaries [VS91] [2]; lingually, jaw with prearticular and 3 coronoids [VS91]; prearticular extends anteriorly to symphysis, overlain by parasymphysial plate anteriorly [VS91]; prearticular forms medial edge of adductor fossa [AC98] [6]; prearticular with anteroventral process forming medial/ventral edge of precoronoid fossa [AC98]; ventral margins of prearticular & infradentaries held together by strip of Meckelian bone [AC98]; prearticular anteroventral process partially separated (might be a separate ossification), and underlies anterior of prearticular [AC98]; parasymphysial plate small, not contacting coronoid [JA97]; Meckelian completely ossified, exposed at symphysis and in all fossae (see image) [AC98] [7]; articular continuous with Meckelian [AC98]; parasymphysial plate with irregularly spaced small teeth [AC98]; dentary with homodont teeth, some specimens with additional smaller outer row [AC98]; dentary with fang pair posteromedial to symphysis [AC98]; coronoids with pits accommodating upper fangs [VS91]; fangs on coronoids 1-2, variably on 3 [AC98]; teeth polyplocodont, with minimal bone of attachment [VS91]; large median gular and large principal lateral) gulars [V80] [SA85]; 7 submandibulars [VS91] [3]; Axial: body narrow & elongate [V80]; vertebral column ossified throughout; centra ring-like (except for ventral slit) near pectorals and caudal fin [V80]; centra made up of large intercentra fused with small pleurocentra [V80] (contra [VS91]: pleurocentra absent); neural arches sutured to dorsal border of intercentra, do not meet or fuse, neural spines paired [VS91]; short, broad dorsal ribs attached (continuously) to arch and intercentrum [V80] [VS91] [5]; enlarged ribs which project ventrally (?!); fin basal scutes absent [V80]; dorsal and anal fins absent or confluent with caudal [V80]; reduced, diphyceral tail [V80]; caudal fin with surrounding finfold [VS91]; Appendicular: dermal shoulder girdle attached to skull through usual series of cleithrum, anocleithrum, supracleithrum & posttemporal [VS91]; clavicles broad & broadly in contact, with small posteromedial interclavicle [VS91]; clethrum with groove to receive ascending process of clavicle [C96]; scapulocoracoid massive, with single, broad attachment to cleithrum [V80] [VS91] [C+02]; probably no scapular blade [C+02]; scapula thickened at attachment to cleithrum [C96]; coracoid plate large, with very large supracoracoid foramen [VS91] [C+02]; glenoid fossa as shallow groove (exact shape unknown) [VS91]; pectoral fins positioned far ventrally [V80] [VS91]; pectorals with 3- mesomeres, with each set specialized, not clearly segmented [S95] [C+02]; pectoral fin radials absent [S95]; humerus elongate (by sarcopterygian standards) [V80]; humerus dorsoventrally flattened, with distinct extensor & flexor surfaces [C96] [C+02] [J+03]; humerus with well-developed deltoid crest [VS91]; entepicondylar postaxial) process absent [VS91] (contra [C+02]: present and dorsoventrally flat); radial condyle ventral [C+02] (contra [S+04]: distally facing) ; radius as long as humerus, with well-developed ventral ridge [VS91]; ulna 50-75% as long as humerus [VS91]; distal forelimb with intermedium and plate of fused radial bones (ulnar plate), not digits [V80]; no distal radials on plate ("ulnare") [C+02]; forelimbs with numerous lepidotrichia; lepidotrichia branched [VS91] pelvics small, set far posteriorly, near caudal fin [V80] [VS91]; Integument: scales small & without cosmine [VS91]; ornamentation on scales tuberculate & ridged [VS91].
Notes: [1] See image at dermintermedial process. [2] splenial, postsplenial, angular, and surangular [AC98]. [3] [VS91] is a somewhat frustrating paper, as the authors' terminology is slightly inconsistent with conventional usage, with occasional internal inconsistencies. Generally, we have attempted to use the most widely accepted terminology. However, we refuse to worry about why the same bone is referred to as both the "submandibulobranchiostegal" and the "preoperculosubmandibular," what that might translate into, or whether this is some bizarre osteolepiform in-joke. [4] As Clack [C02] points out, Panderichthys shows not only tetrapod-like interdigitating sutures in the dermal skull, but also development of other suture types: scarf & tongue in groove. [5] Clack [C02] notes that the ribs are not well known and are "difficult to interpret." [6] [CA98] use the confusing British nomenclature, not followed here. As a reminder: medial means "toward the center line of the body," while (or whilst, if you absolutely must) mesial means "towards the symphysis." [7] Our image erroneously fails to show the Meckelian visible in the precoronoid fossa. The original in [AC98] shows this feature correctly.
Images: The image of the ventral skull has been modified to include a median gular which is not in the original figure.
References: Ahlberg & Clack (1998) [AC98]; Ahlberg et al. (1996) [A+96]; Clack (2002) [C02], Cloutier & Ahlberg (1996) [CA96], Coates 1996) [C96]; Coates et al. 2002) [C+02], Johanson & Ahlberg (1997) [JA97], Johanson et al. (2003) [J+03], Schultze & Arsenault (1985) [SA85], Shubin 1995) [S95], Shubin et al. (2004) [S+04]; Vorobyeva 1980) [V80], Vorobyeva & Schultze (1991) [VS91]. ATW031010.
Elpistostege:E. watsoni Westoll, 1938.
Range: Late Devonian (Givetian? or Frasnian) of North America (Canada)
Phylogeny: Elpistostegalia:: Panderichthys + *.
Characters: snout prominent [VS91]; mouth subterminal [SA85]; premaxilla narrow at symphysis, accommodating unpaired median rostral [SA85]; laterally, premaxilla widens and then tapers under septomaxilla [1] to a narrow suture with maxilla [SA85]; maxilla forms middle section of upper jaw, terminating anteriorly on premaxilla at the level of the lateral rostral [SA85]; snout bones numerous [SA85]; unpaired median postrostral may be present [VS91]; lacrimals uniquely large, broadening anterolaterally [SA85] [VS91]; frontals paired, broad, elongate & meeting parietals anterior to orbit [SA85]; frontals are widest bone in skull roof [SA85]; frontals have interdigitating sutures with postrostrals & nasals [SA85]; prefrontals reaching at least as far anteriorly as frontals [SA85]; prefrontals extensively underlie frontal & lacrimal [SA85]; postfrontal ridge ("eyebrows") [SA85]; parietal meets frontal anterior to orbit [SA85]; parietals narrow between orbits & expanding posteriorly [SA85]; parietal foramen slightly raised [SA85]; parietal extensively underlies postorbital [SA85]; postorbital broadly overlaps parietal [SA85]; parietals & postparietals roughly same anteroposterior length [SA85]; postparietals embayed on posterior margin [SA85]; postorbital broad [SA85]; intertemporal absent [SA85]; postorbital fuses with intertemporal posteriorly [VS91]; orbits located high and closely spaced [VS91]; orbits dorsoventrally elongate and elevated [SA85]; jugal with slight contact or no contact with orbit [SA85]; lower jaw with 3 infradentaries & 3 right, 4 left submandibulars, large median gular [SA85]; 16-17 vertebrae [SA85] [2]; neural arches with smooth, perichondral surface [SA85]; neural arches not fused [C96]; intercentra with endochondral bone internally [SA85]; intercentra forming half-rings, pleurocentra not found [SA85]; scales overlap extensively (~50% as judged by unornamented surface) within scale rows, but only slight overlap between scale rows (i.e. anteroposteriorly) [SA85].
Notes: [1] or the lateral rostral. The septomaxilla probably derives from the lateral rostral, anterior tectal, or both. See, e.g., [C02] (anterior tectal). For simplicity, we use the latter name throughout. [SA85] apparently believe that the septomaxilla derives from the lateral rostral, so it is not clear if their nomenclature is standard. [2] Unclear whether this is the vertebral count for the reconstructed organism, the particular fossil, or both.
References: Clack (2002) [C02]; Coates (1996) [C96]; Schultze & Arsenault (1985) [SA85]; Vorobyeva & Schultze (1991) [VS91]. ATW050415
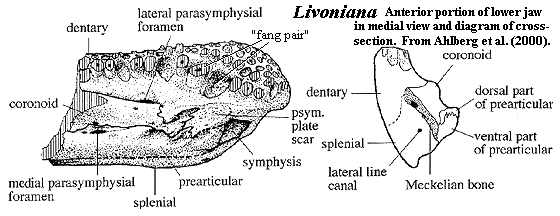 Livoniana: L. multidentata Ahlberg et al., 2000. Known only from 2 partial jaws.
Livoniana: L. multidentata Ahlberg et al., 2000. Known only from 2 partial jaws.
Range: Middle or Late Devonian (Givetian or Frasnian) of the Baltics
Phylogeny: Elpistostegalia:: Elginerpetonidae + ((Metaxygnathus + Ventastega) + Tetrapoda)) + *.
Characters: dentary very robust, expanding into club-shaped structure anteriorly [A+00]; dentary-coronoid suture interdigitating anteriorly, straight posteriorly [A+00]; dentary forms dorsal & anterior margins of symphysial Meckelian exposure [A+00]; parasymphysial plate large & contacts dentary, prearticular & coronoid [A+00]; prearticular forms posterior margin of symphysis [A+00]; anterior coronoid narrow & without teeth [A+00] [1]; precoronoid fossa greatly reduced or absent [A+00]; splenial without extensive medial lamina [A+00]; splenial robust, with deeply-buried lateral line canal [A+00]; splenial dentary suture invisible in lateral view [A+00]; teeth with dentine folding pattern like tetrapods, with no side-branches or enclosed spaces [A+00]; dentary with multiple (5-6) rows of teeth at anterior end [A+00$]; dentary teeth face dorsomedially [A+00]; dentary fang pair within, not medial to, innermost tooth row [A+00$]; dentary fang pair only slightly larger than other teeth [A+00$]; dentary with 3 tooth rows posteriorly, additional rows being added buccally [A+00]; dermal ornament lacks tetrapod "starburst" pattern [A+00].
Notes: [1] Ahlberg et al. [A+00: 537] note that the presence of the lateral & medial parasymphysial foramina (see figure) and the lack of Meckelian exposure between the prearticular & coronoid are "all diagnostic tetrapod features."
References: Ahlberg et al. 2000) [A+00]. ATW050415
checked ATW050624