|
|
Bones: The Braincase |
| The Vertebrates |
The Prootic |
The Prootic
Anatomy
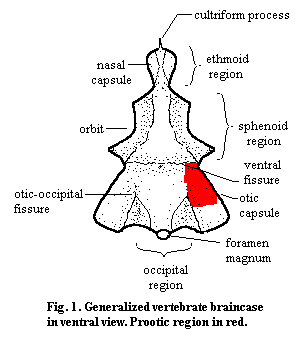
The prootic (in older or European texts, the "proötic") is an endochondral bone of the braincase. It typically forms the anterior and dorsal surface of the otic capsule. The otic capsule contains, in addition to the otic region of the brain itself, the labyrinth and inner ear. The labyrinth is the sensory organ which informs the brain about its orientation and acceleration in a gravity field. As with most endochondral bones bones pre-formed in cartilage) the prootic does not form neat sutures, but tends to merge seamlessly with the other centers of ossification. It co-ossifies with the opisthotic, the other major bone of the otic capsule in many vertebrates, which is usually integrated into the external occipital wall (i.e. the back surface of the skull). The prootic itself may also articulate internally with the occiput, e.g., through the exoccipitals or the supraoccipital. It may also articulate with various dermal bones of the roof or sides of the skull for support, normally the squamosal and parietal. In dinosaurs, for example, the otic capsule can be fixed indirectly by the depression it leaves on the internal surface of the lateral wall of the skull. Most significantly, however, the prootic articulates with the quadrate. In more basal vertebrates, the quadrate simply braces the braincase. In more derived tetrapods*, this articulation forms the fenestra ovalis (fig. 2). In this case, the stapes forms a tight seal with an open area defined by the prootic and opisthotic and transduces airborne sound vibrations to vibrations in the liquid-filled inner ear. The prootic may also bear ventrolateral processes which staple it to the occiput or to one of the sphenoids underlying the braincase, or posterior processes which may reach the external surface of the occipital region.
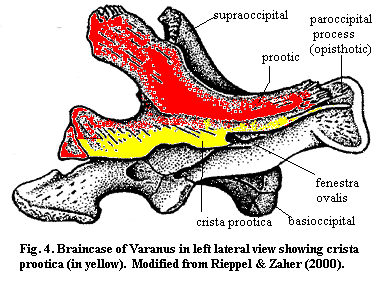 One of the more reliable landmarks on the prootic is the crista prootica. The crista prootica is a ridge or ledge of the prootic which runs antero-posteriorly along the side of the braincase above the otic capsule. Figure
4, modified from Rieppel & Zaher (2000). As shown in the figure, the crista prootica can be elaborated into a major braincase component. In Varanus (the monitor lizard), as in Trilophosaurus and other reptiles, the crista may join with the opisthotics or exoccipitals to form the paroccipital process. This process acts as a sort of horizontal cross-bar across the back of the skull. Anteriorly, the crista may be elaborated into a kind of awning which shelters the foramina from which other cranial nerves communicate with the brain.
One of the more reliable landmarks on the prootic is the crista prootica. The crista prootica is a ridge or ledge of the prootic which runs antero-posteriorly along the side of the braincase above the otic capsule. Figure
4, modified from Rieppel & Zaher (2000). As shown in the figure, the crista prootica can be elaborated into a major braincase component. In Varanus (the monitor lizard), as in Trilophosaurus and other reptiles, the crista may join with the opisthotics or exoccipitals to form the paroccipital process. This process acts as a sort of horizontal cross-bar across the back of the skull. Anteriorly, the crista may be elaborated into a kind of awning which shelters the foramina from which other cranial nerves communicate with the brain.
Functions
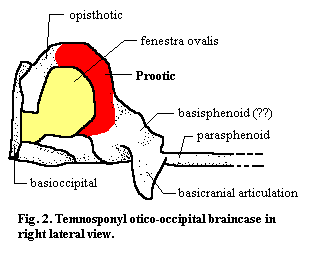
The prootic is associated with the following functions:
(a) Protection of the labyrinth and inner ear: This is perhaps the most ancient and invariable function of the otic capsule generally. The labyrinth consists of the semicircular canals which, in turn have hair cells with small otoliths (roughly, pebbles attached to the hair cells). The canals each have a specific orientation and are so arranged that the movement of the otoliths can be integrated by the brain to determine orientation in a gravity field or acceleration. Since an organ which can detect motion in liquid can detect (on a finer scale to be sure) vibration in liquid, it isn't hard to see how this apparatus might evolve an auditory function. In derived tetrapods, the inner ear contains both the auditory and "balance" sensors. In addition to forming all or part of the otic capsule, the prootic may bear a crista prootica which provides a second layer of protection for the otic capsule and other neural structures.
(b) Lateral support of the braincase: As discussed above, the prootic is a rather protean bone which can attach itself to a variety of supports. See the braincase overview for the importance of lateral support. Again, the crista prootica elaborates on this function by participating in the paroccipital process of the occiput.
(c) Support of/by the stapes: Primitively, the stapes was a rather large bone which provided ventrolateral bracing to braincase through the otic capsule. However, in this position, it naturally carried some external vibrations to the otic capsule. In many early tetrapod lineages, the primary function of the otic capsule became hearing, and the prootic-stapes junction became the fenestra ovalis. In advanced tetrapods*, the stapes loses its support function altogether and becomes a minute element specialized for sound transduction. Thus, the respective roles of the two bones are reversed, with the prootic serving to support the stapes as a sound transducer, rather than being supported by the stapes.
Phylogeny

Osteostracans have a distinct vestibular chamber, but no otic capsule or prootic as such. The placoderms and chondrichthyans both show at least some capsular protuberance of the braincase, but the braincase is a single, undivided mass, whether or not ossified. Interestingly, even at this stage, the braincase frequently bears a shelf or ridge over the otic capsule, so that homologues of the crista prootica may predate the prootic itself. See, e.g., Coates & Sequeira 1998).
Acanthodians and most sarcopterygians are believed to have had a single and undivided otico-occipital ossification. Teleosts have a prootic ossification, but (like a number of teleost bones), it is not clear that this is homologous with the tetrapod bone of the same name. Fig. 3. The teleost prootic may be integrated with the lateral dermal bone through the dermopteric and pterotic, bones which have no straightforward analogs in land animals. The basal tetrapods possessed a recognizable prootic ossification, more or less similar to that shown in Figure 2. One, somewhat aberrant and interesting example (the frog), may be seen at Prootic Bone. Note how the whole otic complex has been oriented laterally and suspended between the squamosals and the pterygoids.
In amniotes, the enlarged supraoccipital links the occiput with the otic capsule, perhaps freeing the stapes to evolve further in the direction of a purely sensory transducer. In cryptodire turtles, the prootic also assumes a special role in supporting and redirecting the force of the major jaw adductor muscles. Among cynodont synapsids, the jaw joint moved anteriorly, and the jaw muscles became associated with the external surface of the dermal bones and in the "new" masseter muscles inside the zygomatic arch. This removed a good many constraints on the size of the brain and the position of the otic capsule. In this lineage, the prootic becomes a much smaller, peripheral element closely associated with the squamosal, eventually merging with the opisthotic to form the petrosal in early mammals.
ATW 010407.
Figure 3 from Mike Oliver's Malawi Cichlids site, and originally from Oliver, MK (1984), Systematics of African cichlid fishes: Determination of the most primitive taxon, and studies on the haplochromines of Lake Malawi (Teleostei: Cichlidae). Unpublished Ph.D. dissertation, Yale Univ. 326+ pp. © 1984 by Dr. Oliver and reproduced here by permission.
checked ATW050507