Sarcopterygii: Elpistostegalia
Elginerpetonidae: Elginerpeton
Taxa on This Page
- Elginerpeton X
- Obruchevichthys X
Elginerpeton: Getting Off on the Wrong Foot
 Elginerpeton has been troublesome. In many ways it looks the way it ought to look for something branching off the sarcopterygian stem shortly before Tetrapoda. However, in some details it has been "a prime source of noise in the data set" [C+02], see also [AL04]. Much of the problem has been associated with the isolated humerus ascribed to Elginerpeton. This is an odd-looking thing with a very small articular head, an almost non-existent shaft, a huge entepicondyle, and no remaining trace of either the ventral ridge or ulnar facet [A98]. We promised we would not take sides in this dispute [A04] vs. [C+04]), and we won't. However, we need to point out that it exists, and that this humerus is controversial.
Elginerpeton has been troublesome. In many ways it looks the way it ought to look for something branching off the sarcopterygian stem shortly before Tetrapoda. However, in some details it has been "a prime source of noise in the data set" [C+02], see also [AL04]. Much of the problem has been associated with the isolated humerus ascribed to Elginerpeton. This is an odd-looking thing with a very small articular head, an almost non-existent shaft, a huge entepicondyle, and no remaining trace of either the ventral ridge or ulnar facet [A98]. We promised we would not take sides in this dispute [A04] vs. [C+04]), and we won't. However, we need to point out that it exists, and that this humerus is controversial.
Fortunately, there's other material to work with. Elginerpeton was a large (1.5m or so), robust, proto-tetrapod which lived somewhere in Scotland in the late Frasnian, a bit under 380 million years ago, a few hundred km northwest of its Latvian cousin, Obruchevichthys. Likely, Elginerpeton inhabited large lakes and freshwater swamplands. There seem to have been a lot of both in the watershed of the mountains of Western Baltica. Elginerpeton had enormously long jaws in its long, narrow skull. The jaws were tubular, particularly anteriorly, and strong for their length. These jaws had the beginnings of all kinds of characteristics that, for one reason or another, became the norm in early tetrapods. It had, for example, coronoid fangs offset from the main tooth row. The precoronoid fossa, a hole in the jaw in front of the first coronoid, is absent; but a foramen has opened near the parasymphysial plates, which have developed their own teeth. These are all tetrapod characters. We don't know much about the rest of the skull, except that it was flattish, bore paired nasal bones (again like a tetrapod), but that the eyes were probably not as large as we might expect of a tetrapod.
The postcranial skeleton is much harder to piece together, as it comes to us in fragments. Several parts of Ahlberg's paper on the subject [A98] have a Cuvier-like magical quality to them. In particular, his reconstruction of almost the entire pelvic girdle from a little nubbin of broken bone is like watching a magician pull a living temnospondyl out of a hat. One is tempted to gasp and applaud, even when he explains, very clearly, just how the trick is done. Its an incredible performance, particularly since the Zoological Journal of the Linnean Society doesn't permit one to use bikini-clad "assistants," deceptive lighting, and similar distractions. The section on the femur is almost as good. However, there is a limit to what human reason can do with one badly abraded example of the middle one third of a single femur.
What's there is odd enough. The femur is distinctly bent through an angle. If the head of the femur is blunt and blade-like, and the leg is held straight out, as expected in most Devonian tetrapods, the tibia would project anteroventrally and the bottom of the "foot" (if any) would face posterodorsally. Note that the reconstruction omits any conjectural trochanters on the femur. This is because, as Ahlberg notes, their predicted positions make no sense, based on the orientation of the adductor blade. One possible conclusion is that Elginerpeton didn't have digits. But this would still leave the lower leg hanging at a really peculiar angle. And, speaking of leaving things hanging, we are going to have to leave this issue hanging until we have had a chance to look in more detail at the toes of Acanthostega. ATW050430.
Obruchevichthys: "... and so do his cousins whom he reckons up by dozens"
 Obruchevichthys gracilis obviously doesn't amount to much by itself. It consists of two overlapping specimens of anterior jaw which probably, despite the name, come from two different species [AC98]. Both look like cheap imitations of Elginerpeton. That is, they lack some of the stylistic refinements of the Scottish product, such as small coronoid fangs, smooth ventral splenial surface, or an accessory dentary tooth row. Obruchevichthys has a blocky, industrial-grade, look to it -- a sort of blue-collar Elginerpeton.
Obruchevichthys gracilis obviously doesn't amount to much by itself. It consists of two overlapping specimens of anterior jaw which probably, despite the name, come from two different species [AC98]. Both look like cheap imitations of Elginerpeton. That is, they lack some of the stylistic refinements of the Scottish product, such as small coronoid fangs, smooth ventral splenial surface, or an accessory dentary tooth row. Obruchevichthys has a blocky, industrial-grade, look to it -- a sort of blue-collar Elginerpeton.
The real significance of Obruchevichthys lies in what it is not. Specifically, it is not Elginerpeton. What this suggests is that Late Devonian tetrapods were not weird one-offs, evolving only in peculiar environments or from isolated branches of phylospace. They were likely a more diverse bunch, with family all over, and active in a number of different ecological guilds. This may be an important point. Writers have had a tendency to posit environments for tetrapod evolution which are unusual. Obruchevichthys suggests that, whatever environmental conditions were driving tetrapod evolution in the Devonian, these conditions were relatively common.
Common -- but perhaps not yet universal. It remains possible that the tetrapod stem group was originally limited to Baltica. Elginerpeton and Obruchevichthys are both from Baltica, and they remain the only well-established Frasnian members of this grade of almost-tetrapod. In fact, all of the Frasnian Elpistostegalia are probably from Baltica. Even when we include trackways, the result is the same. The Australian Genoa River trackway was historically regarded as Frasnian, but more recent evaluation suggests that it was made substantially later [C97]. All others are either from Baltica or date from the Famennian.
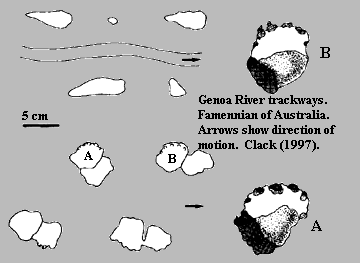 We do have to waffle on this generalization, because some workers place the Australian Metaxygnathus just below the Frasnian-Famennian boundary [LG04]. Further, the entire world-wide diversity of known Devonian tetrapods and their near kin is still quite small -- perhaps a dozen species as of this writing. While the numbers are growing, the sample size is still very small. However, even if the Frasnian tetrapods were confined to Baltica, Baltica is still a very large area.
We do have to waffle on this generalization, because some workers place the Australian Metaxygnathus just below the Frasnian-Famennian boundary [LG04]. Further, the entire world-wide diversity of known Devonian tetrapods and their near kin is still quite small -- perhaps a dozen species as of this writing. While the numbers are growing, the sample size is still very small. However, even if the Frasnian tetrapods were confined to Baltica, Baltica is still a very large area.
So, what kind of area was it? Mostly it was very wet. Sea levels were very high up until the "Kelwasser Event" at the very end of the Frasnian. We also know that the Acadian Orogeny had been building tall mountains in Eastern Laurentia (North America and Greenland) near where Baltica and Laurentia were stapled together. This had been going on since the Pridoli. The interior of Laurentia was quite arid. The natural implication is that Baltica was receiving all the rain from the soggy Trade Winds blowing inshore from the Old Tethys Sea, while most of Laurentia was in the rain shadow of the Acadian Range. At just about this time, land plants were beginning to push their way inland, developing stronger woody stems and more substantial root systems. Insects were not yet a major factor, but numerous other arthropods were adapting to these new, plant-dominated environments. Baltica was slightly north of the equator. It is generally supposed that the climate was warm, as well as wet. Frankly, we have some lingering doubts about the conventional wisdom on this point. Baltica was a big place. The temperature may have varied considerably -- but it was certainly wet.
Now, at this point, you are probably wondering, "Enough with the weather report, already! What's this got to do with tetrapods? Especially frankly aquatic ones like Obruchevichthys -- its not like they needed galoshes and an umbrella!" But such impertinence will gain you nothing. It is beneath our dignity to reply to such drivel. Indeed, it will be necessary to wander yet further afield before we can get back to the point. Besides, we have a few more jaw taxa to finish up first, so we need to drag this out a bit longer.
Image: a Late Devonian estuarine gallery forest in Baltica from the New Brunswick Museum. This is very much the sort of setting in which the remains of Elginerpeton were deposited. ATW050501.

Elginerpeton: E. pancheni Ahlberg, 1995. Incomplete jaws, premaxilla, postorbital and other cranial fragments; some isolated postcranial material, including one half neural arch, dorsal portions of pectoral and pelvic girdles, a humerus, and femoral and tibial fragments.
Range: later Frasnian of Scotland [A95]
Phylogeny: Elginerpetonidae : Obruchevichthys + *.
Characters: up to 1.5 or possibly more (skull ~40 cm) [A98]; skull:skull long & triangular[A95]; snout long and laterally compressed [A95]; single median rostral as in Ichthyostega [A95]; premaxilla with long posterior extension [A95]; premaxilla with extended attachment areas for nasal and vomer [A95]; irregular, radiating ornament on premaxilla & infradentaries [A95];nasals large [C96]; orbit large [A98]; postorbital anteroposteriorly deep [A98]; palate: vomer with fangs [A95]; jaw:lower jaw long and very slender [A95]; convexity of dentary more conspicuous than in Obruchevichthys [A95]; but jaw transversely thin posteriorly [A+05]; dentary reaches almost to glenoid as in tetrapods [A+05]; bones of lateral face closely interdigitated or partially fused [AC98]; ventral margins of splenial & anterior postsplenial thickened & rounded [AC98]; suture between dentary & splenials with marked groove [AC98]; parasymphysial plate and foramina present [A95]; parasymphysial plate with paired foramina posterolateral to tooth row and large lateral parasymphysial foramen [A95]; prearticular extends almost to parasymphysial foramina [AC98]; prearticular with denticulated dorsal ridge & very narrow ventral lamina, leaving large Meckelian fossa [AC98]; ventral margins of prearticular & infradentaries held 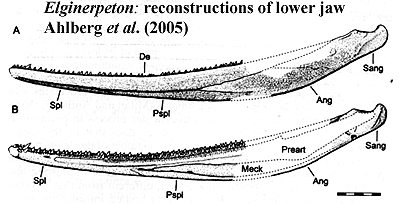 together by strip of Meckelian bone [AC98]; "absence of exposed Meckelian bone dorsal to the prearticular" [A95] [1]; Meckelian ossified [AC98]; coronoids with tall distinct vertical laminae [AC98]; surangular forming outer face and posterior margin proximal jaw area [A+05]; surangular tightly fused to articular [A+05]; surangular forming rounded retroarticular process [A+05]; posteroventral area of outer surangular depressed as in Ventastega, Ichthyostega & Densignathus [A+05]; surangular, dorsal concavity anterior to preglenoid process of articular [A+05$]; articular exposed laterally in low, dorsal preglenoid process anterior to glenoid, with 3 foramina for ramus mentalis externus of cranial nerve VII [A+05]; articular separating prearticular from surangular so that prearticular fails to contact angular (as in Densignathus and Metaxygnathus), resulting in long, continuous medial exposure of Meckelian [A+05]; angular, medial lamina absent [A+05]; articular grades smoothly onto floor of adductor fossa [A+05]; dentition: parasymphysial plate with tooth row & fangs [A95]; coronoid fangs small, esp. in larger specimens [A95]; auxiliary tooth row on dentary [A95]; anterior coronoid, with fang pair (same size as marginal teeth) lingual to main tooth row [A98] [6]; accessory tooth row on dentary [A95], [A95]; axial: vertebrae with zygapophyses present, but weakly developed and without joint-like, unfinished bone surfaces or lateral expansion [A98]; transverse processes absent [A98]; neural arches low [A98$]; neural arches with roughened medial surface for synchondrosis, but not fusion, with antimeres [A98]; neural arch with external attachments for supraneural ligament, not canal [A98] [C96$]; appendicular: postbranchial lamina of cleithrum probably absent [A98]; cleithrum wraps around scapulocoracoid ventrally, with furrow [2] at the join [A98]; cleithrum does not merge with scapulocoracoid [A98]; scapular blade absent [A98]; massive posterior infraglenoid buttress [A98]; deep subscapular fossa [A98]; humerus dorsoventrally flattened [A04]; humeral head small & proximal region short [A04]; transverse (ventral) humeral ridge absent [A04]; prominent pectoral process [A04]; supinator process present, close to ectepicondyle [A04] [3]; ectepicondyle tall & sharply ridged [A04]; ectepicondylar foramen absent [A04]; entepicondyle thin, flat, with rounded end & continuous with body of humerus [A04]; radial facet elongate, anteroventrally positioned [A04]; iliac process present [A98]; post-iliac process short, with
together by strip of Meckelian bone [AC98]; "absence of exposed Meckelian bone dorsal to the prearticular" [A95] [1]; Meckelian ossified [AC98]; coronoids with tall distinct vertical laminae [AC98]; surangular forming outer face and posterior margin proximal jaw area [A+05]; surangular tightly fused to articular [A+05]; surangular forming rounded retroarticular process [A+05]; posteroventral area of outer surangular depressed as in Ventastega, Ichthyostega & Densignathus [A+05]; surangular, dorsal concavity anterior to preglenoid process of articular [A+05$]; articular exposed laterally in low, dorsal preglenoid process anterior to glenoid, with 3 foramina for ramus mentalis externus of cranial nerve VII [A+05]; articular separating prearticular from surangular so that prearticular fails to contact angular (as in Densignathus and Metaxygnathus), resulting in long, continuous medial exposure of Meckelian [A+05]; angular, medial lamina absent [A+05]; articular grades smoothly onto floor of adductor fossa [A+05]; dentition: parasymphysial plate with tooth row & fangs [A95]; coronoid fangs small, esp. in larger specimens [A95]; auxiliary tooth row on dentary [A95]; anterior coronoid, with fang pair (same size as marginal teeth) lingual to main tooth row [A98] [6]; accessory tooth row on dentary [A95], [A95]; axial: vertebrae with zygapophyses present, but weakly developed and without joint-like, unfinished bone surfaces or lateral expansion [A98]; transverse processes absent [A98]; neural arches low [A98$]; neural arches with roughened medial surface for synchondrosis, but not fusion, with antimeres [A98]; neural arch with external attachments for supraneural ligament, not canal [A98] [C96$]; appendicular: postbranchial lamina of cleithrum probably absent [A98]; cleithrum wraps around scapulocoracoid ventrally, with furrow [2] at the join [A98]; cleithrum does not merge with scapulocoracoid [A98]; scapular blade absent [A98]; massive posterior infraglenoid buttress [A98]; deep subscapular fossa [A98]; humerus dorsoventrally flattened [A04]; humeral head small & proximal region short [A04]; transverse (ventral) humeral ridge absent [A04]; prominent pectoral process [A04]; supinator process present, close to ectepicondyle [A04] [3]; ectepicondyle tall & sharply ridged [A04]; ectepicondylar foramen absent [A04]; entepicondyle thin, flat, with rounded end & continuous with body of humerus [A04]; radial facet elongate, anteroventrally positioned [A04]; iliac process present [A98]; post-iliac process short, with 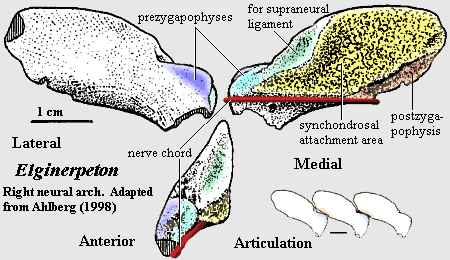 horizontal orientation [A98]; post iliac process laterally projecting [A98$]; iliac neck placed anteriorly, above anterior end of acetabulum [A98], with iliac neck with foramen [A98] [C96$]; femur with intercondylar fossa, adductor blade, & adductor crest [A98]; femur adductor blade with regular transverse grooves ("fluted") [A98]; femur with strong tortion, intercondylar fossa facing anteroventrally [A98] [5]; partial crest of fibular condyle sharper & more prominent than fibular condyle [A98]; tibia short & blocky, with moderate anterior cnemial crest [A98]; tibial facet for tibiale extends further toward flexor surface than facet for intermedium [A98] [4]; typical stem tetrapod ornament of tubercles and short ridges [A+05]; unmodified basal lateral line system [AC98].
horizontal orientation [A98]; post iliac process laterally projecting [A98$]; iliac neck placed anteriorly, above anterior end of acetabulum [A98], with iliac neck with foramen [A98] [C96$]; femur with intercondylar fossa, adductor blade, & adductor crest [A98]; femur adductor blade with regular transverse grooves ("fluted") [A98]; femur with strong tortion, intercondylar fossa facing anteroventrally [A98] [5]; partial crest of fibular condyle sharper & more prominent than fibular condyle [A98]; tibia short & blocky, with moderate anterior cnemial crest [A98]; tibial facet for tibiale extends further toward flexor surface than facet for intermedium [A98] [4]; typical stem tetrapod ornament of tubercles and short ridges [A+05]; unmodified basal lateral line system [AC98].
Image: Elginerpeton life reconstruction by Prof. Per Ahlberg, with completely unauthorized digital tinkering by ATW. It is dated 1993 and does not include some important reinterpretations of the limbs contained in [A98].
Notes: [1] The Meckelian, so far as we know, is never dorsal to the prearticular. The quoted character presumably means that the prearticular does not cover the the Meckelian foramen, so that the Meckelian bone is exposed in medial view, as explained in [A+05]. [2] See discussion of this "groove" at anterior ridge. [3] [A98] erroneously states that the supinator process is close to the "entepicondyle." However, the figure in [A98] shows the correct relation. [4] "this joint morphology [may be] related to hyperflexion of the leading edge of the foot." [A98: 124]. [5] as [A98] notes, when this is combined with the inferred position of the fibula dorsal & somewhat anterior to tibia, the implication is that the flexor surface, and the lower leg, were oriented not only posteriorly, but posterodorsally! In other words, with the leg stuck out at right angles, the sole of the feet would face partially upwards. This accounts in part for the odd orientation of the leg in Ahlberg's life reconstruction -- which is much less aggressively interpreted than his written description. [6] an image of this area of the jaw, with some commentary, can be found at Elginerpetonidae.
References: Ahlberg (1995) [A95]; Ahlberg (1998) [A98]; Ahlberg (2004) [A04]; Ahlberg & Clack (1998) [AC98]; Ahlberg & Luksevics (2004) [AL04]; Ahlberg et al. (2005) [A+05]; Coates (1996) [C96]; Coates & Clack (1991) [CC91]; Coates et al. (2002) [C+02]; Coates et al. 2004) [C+04]; Daeschler et al. (1994) [D+94]; Jarvik 1996) [J96]; Lebedev & Coates (1995) [LC95]. ATW051113.
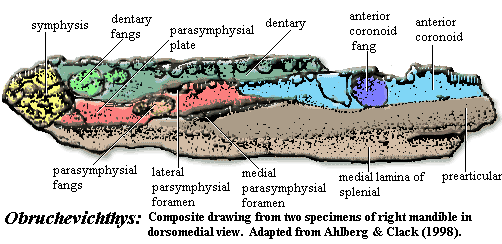
Obruchevichthys: O. gracilis Vorobyeva, 1977. Lower jaw. The two jaw fragments assigned to this species differ in some details and probably represent two different species [AC98].
Range: Late Frasnian of Latvia & Russia.
Phylogeny: Elginerpetonidae : Elginerpeton + *.
Characters: "inflated" dentary and other characters of Elginerpetonidae [A95]; "inflation" of dentary not as marked as in Elginerpeton [A95]; splenial with sharp ventral edge [AC98]; dentary accessory teeth absent [AC98]; fangs proportionately larger than Elginerpeton [A95].
References: Ahlberg (1995) [A95]; Ahlberg & Clack (1998) [AC98]; Clack (1997) [C97]; Long & Gordon (2004) [LG04].
