 A Lot of Rot: the World of the Tetrapods
A Lot of Rot: the World of the Tetrapods| Tetrapoda | ||
| The Vertebrates | Tetrapoda |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Sarcopterygii
│
└─Tetrapoda
├─Acanthostega
└─┬─Ichthyostega
└─┬─Tulerpeton
└─┬─Crassigyrinus
└─┬─Colosteidae
│ └─Greererpeton
└─┬─Baphetidae
└─Tetrapoda*
├─Temnospondyli
└─┬─Lepospondyli
└─Reptiliomorpha
|
Contents
Overview |
A Lot of Rot: the World of the TetrapodsWe have hinted in various other places that rot may be the key to Late Devonian and Mississippian (Early Carboniferous) environments. It is now time to develop that thesis, since we will never understand the evolution of tetrapods without understanding the environment in which they evolved. It's a very long story, full of chemistry and mostly concerned with the evolution of plant tissues and fungi. The prospect of explaining this stuff is so utterly discouraging that we feel compelled to recommend that you skip the whole thing. Furthermore, we expressly disclaim any and all liability for nausea, vomiting, cerebral atrophy, sleep disturbances, uncontrollable ophthalmic tremors, progressive neuronal putrification, acute mental atrophy, or any other physical or psychiatric side effects which readers may experience. You have been warned.
 What do we know about the environments at the actual sites in which Devonian tetrapods are found? Not a great deal, as it turns out. If we ignore Tulerpeton, it makes things more comprehensible. Tulerpeton was found in what is supposed to be a completely marine environment, but with other possibly terrestrial or near-shore remains. Lebedev & Clack (1993); Lebedev & Coates (1995). Its rather hard to explain, except as a rafting accident, or some other weird one-off, and we won't even try to make sense of it.
What do we know about the environments at the actual sites in which Devonian tetrapods are found? Not a great deal, as it turns out. If we ignore Tulerpeton, it makes things more comprehensible. Tulerpeton was found in what is supposed to be a completely marine environment, but with other possibly terrestrial or near-shore remains. Lebedev & Clack (1993); Lebedev & Coates (1995). Its rather hard to explain, except as a rafting accident, or some other weird one-off, and we won't even try to make sense of it.
The others, to the extent anything is known about their depositional environment at all, share two features. They are all associated with meandering rivers, and they come with salad. That is, they are also associated with plant material (usually unidentifiable). Densignathus and Hynerpeton: Red Hill, usually interpreted as an oxbow lake near a meandering river, with shrub-like lycopsids and Archaeopteris trees in the neighborhood. Daeschler et al. (1994); Clack (1997); Daeschler (2000); Shubin et al. 2004). Ichthyostega and Acanthostega: Aina Dal and Britta Dal Formations with meandering rivers, point bars and unidentified plants. Coates 1996); Clack (1997); Blom (2005). Genoa River tracksite with meandering streams. Clack (1997). Sinolepis: plant materials, nearshore or fresh water vertebrate remains, possibly deltaic. Zhu et al. 2002).
The existence of meandering rivers is significant. In the absence of well-established terrestrial plants, rivers don't form gently curved meandering river beds. They cut deep, narrow channels or spread out into a maze of braided streams. The implication of meandering rivers is that land plants weren't just a marginal element of these systems. By the Late Devonian, land plants covered substantial areas,  and tetrapods lived in the waters in or near these areas.
and tetrapods lived in the waters in or near these areas.
The identities of the plants themselves are reasonably known only at the latest Famennian Red Hill tetrapod site. As mentioned, this site is generally interpreted as an ox bow lake near a large river. Here, fragments of Archaeopteris are plentiful; and we can be certain that the lake, like much of the Devonian landscape, was surrounded by a forest of these huge seed ferns progymnosperms). Shear & Selden 2001). Archaeopteris was a true tree, with a woody trunk, xylem, secondary cambium, and leaves. DuBuisson et al. (2002). See images and additional discussion at Devonian, Late Devonian and Frasnian. Below the Archaeopteris canopy, we would see a ground cover of Protolepidodendron up to 30 cm in height), with stands of various lycopsids. Shear & Selden (2001). The shallows of the  lake itself were choked with dense, bushy Rhacophyton, growing as tall as 2 m. Additional information on the flora of Red Hill (and much else besides) can be found at Dennis Murphy's Devonian Times site.
lake itself were choked with dense, bushy Rhacophyton, growing as tall as 2 m. Additional information on the flora of Red Hill (and much else besides) can be found at Dennis Murphy's Devonian Times site.
For our purposes, we should also note that Red Hill, and all other tetrapod sites about which we have detailed knowledge, shows signs of periodic flooding -- perhaps seasonal, perhaps not. The taphonomy is also important. Plant remains are found as dense mats of poorly (or only secondarily) oxidized litter. Shear & Selden 2001). It seems likely that these two observations are connected, although it is not a certainty. But, whether the forest litter was washed into the lake or accumulated in some other way, it seems almost certain that the lake was weed-choked and generally anoxic.
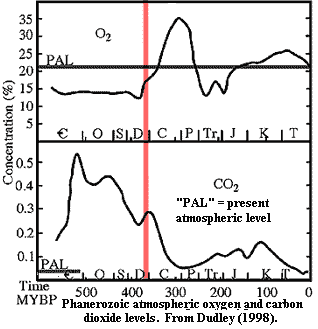 While these shallow water environments may have been low in oxygen, the air above them was not. On the right, we show the usual Phanerozoic oxygen and carbon dioxide trends. The graph is from Dudley 1998), but it is largely based on the work of Robert Berner of Yale and his colleagues. As a preliminary matter, note that atmospheric carbon dioxide levels begin to drop long before oxygen begins to accumulate. This is a bit hard to explain. Fortunately, more recent studies tell a more reasonable tale of a modest, gradual decline in carbon dioxide levels from the beginning of the Cambrian, followed by an abrupt and precipitous fall beginning in the Frasnian or thereabouts. Berner & Kothavala (2001). As an independent check on the overall oxygen trend, it is notable that fusain is found for the first time in the Late Devonian. This strongly implies that (a) oxygen levels had reached the point at which fires could occur (~15%) [1]and (b) that there were plants to burn. Berner et al. (2003).
While these shallow water environments may have been low in oxygen, the air above them was not. On the right, we show the usual Phanerozoic oxygen and carbon dioxide trends. The graph is from Dudley 1998), but it is largely based on the work of Robert Berner of Yale and his colleagues. As a preliminary matter, note that atmospheric carbon dioxide levels begin to drop long before oxygen begins to accumulate. This is a bit hard to explain. Fortunately, more recent studies tell a more reasonable tale of a modest, gradual decline in carbon dioxide levels from the beginning of the Cambrian, followed by an abrupt and precipitous fall beginning in the Frasnian or thereabouts. Berner & Kothavala (2001). As an independent check on the overall oxygen trend, it is notable that fusain is found for the first time in the Late Devonian. This strongly implies that (a) oxygen levels had reached the point at which fires could occur (~15%) [1]and (b) that there were plants to burn. Berner et al. (2003).
No one seems to doubt that the spread of terrestrial plant life in the Late Devonian was the principal cause of the rise in atmospheric oxygen. But why the preceding gradual reduction, and why the concomitant free fall in carbon dioxide? That, in fact, is what this essay is all about. Our thesis, somewhat over-simply stated, is that the evolution of wood resulted in a runaway positive feedback loop towards the end of the Devonian. The problem was that plants are very good at making wood, and they do it even better when oxygen levels are high. However, recycling wood is an extraordinarily difficult, dangerous, and slow job. Wood was, in fact, the plastic waste of the Late Paleozoic: indigestible, often toxic, physically dangerous and sometimes environmentally devastating. It took almost 50 My for a group of fungi, the Basidiomycota, to evolve the complex array of metabolic tools necessary to save the planet from -- yes -- tree pollution.
This speculation has also been advanced by Dudley 1998), and probably by others. Our particular take on it still leaves the matter as a speculation. However, we believe that there is much more to say on the topic, particularly about the biochemistry which drove this near-catastrophe and its environmental effects -- effects which, in turn, seem to have driven the evolution of the early tetrapods. The biochemical foundations of the crisis were laid deep in Archean time, and that is where we will pick up the tale on the next page. ATW050624.
GO TO PART 2. Hazardous Waste & Inefficiency: A Short History of Rubisco
[1] One might suppose that the critical parameter is pO2, the partial pressure of oxygen, not the mole percent O2. For reasons which are entirely unclear to us, all writers on Paleozoic atmosphere assume (a) that total atmospheric pressure was constant throughout the Phanerozoic and (b) that pN2, the partial pressure of nitrogen, was also approximately constant. We can think of no reason why this would necessarily be the case, and several reasons why it might not be. However, our sample of the literature was small, and we leave the problem for another day.
 |
|
The temnospondyl Eryops lunges after the shark Orthacanthus. Another, smaller temnospondyl (Trimerorhachis) swims in the foreground. Painting © Doug Henderson, reproduced with permission |
Tetrapoda = Stegocephalia)
from the Late Devonian (Famennian).
Elpistostegalia::: (Metaxygnathus + Ventastega) + * : Acanthostega + (Ichthyostega + Sinostega + (Densignathus + (Hynerpeton + (Tulerpeton + (Ossinodus + (Whatcheeriidae + (Crassigyrinus + (Colosteidae + (Spathicephalus + (Baphetidae + Tetrapoda*))))))))))).
Dermal skull solidifies, loses complex kinesis and becomes more closely associated with braincase; sphenethmoid and otic-occipital units of braincase become more integrated; snout is longer, post-orbital region shorter; palate changes little; palatoquadrate gradually reduced; supratemporal stiffens former parietal-postparietal hinge; gills and opercular series (including extrascapulars) lost; dermal skull not attached to pectoral girdle; primitively, notochord still prominent and extends into midbrain; atlas-axis complex originally absent & evolved convergently in several later lineages; primitively, retain large, crescent-shaped intercentra connected to neural arches by short ribs, small paired pleurocentra; zygapophyses on vertebrae; cleithrum freed from scapulocoracoid; more-or-less fused sacrum with sacral rib; pelvic girdle from three, enlarged centers and sutured together; iliac blade attached to vertebral column; glenoid and acetabulum significantly reoriented to face ventrolaterally; limbs with well-defined joints, carpals, tarsals & digits; dermal fin rays lost. Stem tetrapods now believed to be aquatic based on groove for aortic arch on ceratobranchials in Acanthostega.
Links: Introduction to the Tetrapoda; Terrestrial vertebrates; Definition of Tetrapoda; Fossil tetrapods; Tetrapoda -- The Dinosauricon; Lecture 8 - Tetrapods; Literature - Tetrapoda; Linking fish to the land; Devonian Times - More about Lobe-Fin Fishes; Re- tetrapoda. ATW030127.
checked ATW040813