Quick quizz. Which of these is an amphibian?



| Tetrapoda | ||
| The Vertebrates | Amphibians, Systematics, and Cladistics |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Sarcopterygii
│
└─Tetrapoda
├─Ichthyostega
└─┬─Tulerpeton
└─┬─Crassigyrinus
└─┬─Colosteidae
└─┬─Baphetidae
└─Tetrapoda*
├─Temnospondyli
└─┬─Lepospondyli
└─Reptiliomorpha
|
Contents
Overview |
| Quick quizz. Which of these is an amphibian? |
|
This unit begins with not one but two taxonomic controversies. The first of these (see quizz, right) comes about through a clash of paradigms (scientific methodologies and perspectives) and the other through how best to define things within a single methodology. Each revolve around a question, the first, what is an amphibian? and the second (considered on the following page), what is a tetrapod? If you find such taxonomic quibbling unintersting (and if you do we certainly don't blame you) feel free to skip ahead to Ichthyostega, where we return to a consideration of the procession of life through deep time (via Beowulf)
Hello to those folks still here. Now, in any biology textbook, Amphibians, Class Amphibia, are generally defined as vertebrate animals that lay their eggs in water and undergo a metamorphosis from a juvenile water-breathing tadpole or larval aquatic form to an adult air-breathing terrestrial form. They include frogs and toads, newts and salamanders, and little known animals called caecilians, limbless amphibians that resemble snakes or large earth worms), as well as a number of extinct palaeozic forms, labyrinthodonts and lepospndyls. Simple? Well, as it turns out, not really.
Like all such statements, this is something of a generalisation. Some amphibians, like the mexican walking fish (axolotl) retain gills in adulthood, others like the Plethodontid salamanders lay eggs on land that hatch into miniature adults (Carroll 2009, p.200) but still require moist environments.
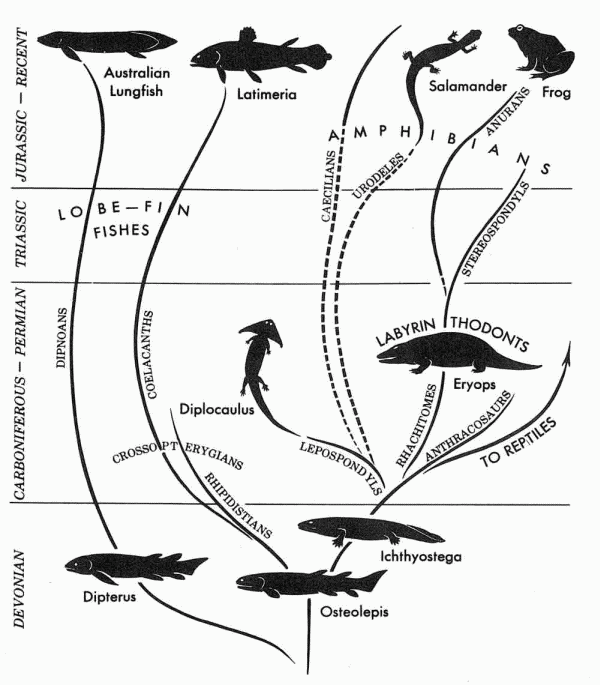 |
| Evolution of lobe-fin fish and amphibians. From E.H. Colbert, Evolution of the Vertebrates, 2nd ed., 1969, copyright John Wiley & Sons. MAK111110 |
 |
| Cross section of Labyrinthodont tooth (from the Triassic genus Mastodonsaurus). from St. George Mivart, F.R.S. On The Genesis of Species London: Macmillan and Co. 1871, public domain, Wikipedia. MAK111110 |
In the first comprehensive systematic and scientific classification of the natural world, the 18th century Systema Naturae of the Swedish botonist Linnaeus, amphibians and reptiles were grouped together in a single category. By the way, defining "Reptiles" is another taxonomic headache, which we've devoted an entire page to it as well. This is because amphibians and reptiles are pretty interesting creatures, and prehistoric amphibians and reptiles more interesting again, especially to palaeo geeks like ourselves. But because of changing paradigms, according to which emphasis switches from one methodology to another, amphibians and reptiles don't mean the same now as they used to. Normally a bit of historical background would be in order. But we'll skip the intro and go straight to the chase. By the start of the 20th century, the main lines of the the evolutionary tree of life had been sketched out. Although many details have changed since then, and will continue to do so, we had a pretty good general idea of which group evolved from which. So in the early to mid 20th century, vertebrate paleontologists such as A.S. Romer (for reptiles) and G.G. Simpson (for mammals) had developed an understanding of vertebrate evolution and phylogeny based on a system that for the sake of convenience we refer to as Evolutionary Systematics (at the time, this term was rarely used). Others such as Raymond Moore developed similar arrangements for invertebrates, and paleobotoanists did the same for plants. The books of these authors can still be read and hold their own as classic reference works today.
The diagram at the right is from Edwin Colbert's Evolution of the Vertebrates (Colbert 1969, p.90), and gives the consensus view of amphibian evolkution in the mid 20th century. This is still very much how we see the relations between the different groups now. The lungfish are a somewhat more distant branch than the rhipidistians like Osteolepis, which were ancestral to the first amphibians like Ichthyostega. From an Ichthyostega-like ancestor, or rather something a bit more advanced, there was a Carboniferous adaptive radiation not only of various Paleozoic amphibians but also of the ancestors of both reptiles and modern amphibians (Lissamphibia). Two main Palaeozoic groups (given, like Lissamphibia, the Linnaean rank of subclass) stand out, the large and diverse Labyrinthodonts (represented by the well known early Permian genus Eryops) and the small Lepospondyls (represented by the distinctive Diplocaulus). The Lepospondyls are a motley assemblage that has nothing in common other than small size and associated loss of primitive characteristics (the fact that they are considered a natural group today is in the opinion of the present author the result of confusing convergences associated with minaturisation (Carroll 2009, pp.155-6) with shared ancestral features). The Labyrinthodonts or "maze teeth", are named after the complex pattern of infolding of the dentin and enamel of the teeth, a characteristic they inherited from their fishy, rhipidistian ancestors (right, below) and basically include all Paleozoic amphibians apart from Leposondyls, and all Mesozoic ones not included under the Lissamphibia. This was a very diverse and indeed vaguely defined group; other than the structure of their teeth they are defined by complex vertebrae (as distinguished from the simple spool-like vertebrae of the leposondyls) and and a soild and heavily armoured and skull roof, another feature inherited from their lobe fin fish ancestors, and in turn inherited by early reptiles (captorhinomorphs and anapsids). This gives them their alternative, 19th century, name, Stegocephalia - "plated heads".
Traditionally (e.g. Romer 1933, 1954, and 1966; Colbert 1969; Carroll 1988) the Labyrinthodonts are divided into three smaller groups or orders. The Devonian Ichthyostegalia are the ancestral forms, in keeping with the evolutionary systematic tendency to posit a short-lived stem group (e.g. cotylosaurs, thecodonts, condylarths...) from which all the other lineages evolved. The Temnospondyli are the main line of labyrinthodont evolution, including both large and small types. They developed progressively more simplified vertebrae through time as they reverted to an increasingly aquatic existence, hence the distinction between the earlier, more terrestrial types, like Eyrops, who belong to the suborder Rachitomi, and the Triassic aquatic capitosaurs and metoposaurs, suborderStereospondyli, in which the vertebrae only consist of two elements., Finally, the Anthracosauria are another diverse group that include both aquatic and terrestrial forms, and were considered somewhat more closely related to reptiles.
Using the word prehistoric in the colloquial sense - before history, before man, millions of years ago, in deep time. For example the present writer as a child a collection of those wonderful How and Why Wonder Books; one of which was called Prehistoric Mammals. In the more technically correct archeological sense however, prehistoric refers to the human period before recorded history, for example the neolithic, or the late palaeolithic. While this is a long time ago, it's not yet in the millions of years (see for example our tabulation of orders of ten gradations of time).
The problem with all these sorts of definitions is that they can be somewhat vague and arbitrary. For example, modern amphibians (Lissamphibia) are probably just as distinct and specialised in relation to the heavy skulled Permo-Carboniferous labyrinthodonts as modern reptiles are. Both these groups evolved from labyrinthodonts, that is an objective phylogenetic fact, but apart from one needing to return to water to lay eggs and the other not, is there a reason why one should be included with them in the same category (Class Amphibia) and the other placed in a different category (Class Reptilia).
The cladistic revolution of the 70s and 80s was an attempt to introduce greater rigour into biological classification. It was pointed out that because the definition of Subclass Labyrinthodontia, and for that matter of class Amphibia, is simply based on primitive characteristics inherited from lobefin fish, and because Labyrinthodonts/Amphibians gave rise to creatures that are not Labyrinthodonts (to wit, Reptiles, Lepospondyls, and Lissamphians) or Amphibians (to wit, Reptiles), neither constitutes a natural group, and therefore in the interests of greater precision such terms - as well as others such as Protista, invertebrates, Reptiles, Thecodonts, Condylarths etc - should be discarded. That is why most vertebrate paleontologists now no longer say amphibian but "tetrapod", the group that includes both amphibians and their descendents; and not reptile but amniote (reptiles and their descendents). They restrict the word "amphibian" to a more limited grouping, either the last common ancestor of all recent amphibians and its descendents, or all animals more closely related to (insert here any recent species of amphibian) than to (insert here any recent species of reptile, mammal, or bird). Creatures that previously were referred to as amphibians but fall outsuide these definitions (some labyrinthodonts for example) are no simply called "tetrapods". On the other hand, zoologists, who deal with recent amphibains (a well recognisable group) happily continue to use the old Linnaean terminology and find no reason to change.
|
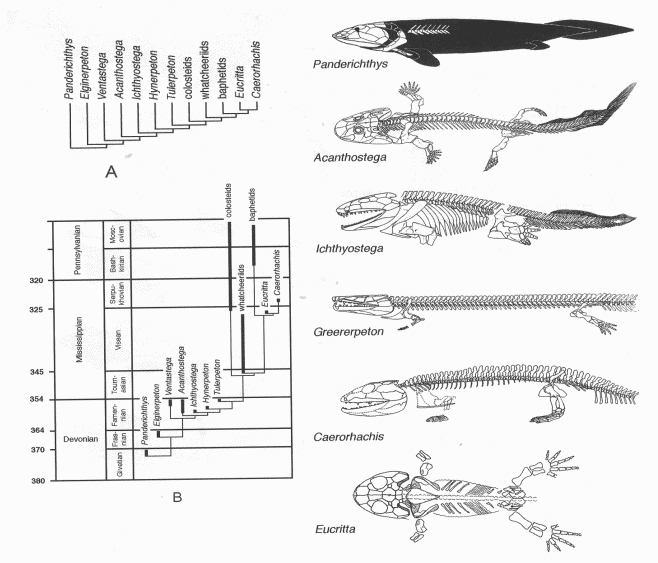
| What is meant by "amphibian"? From classroom handout by Dr. Stuart Sumida. |
For cladistics what is important is not the taxonomic ranking (order, subclass, whatever), or the stratigraphic range in deep time, or which groups evolve into which (ancestors and decsendents), but what characters to use to define a group of organisms, and how choice of characters effects the statistical outcome of the phylogenetic relationships between these different organisms, represented diagramatically as a phylogenetic tree, or cladogram. Thus, with amphibians (or tetrapods) the question is not how did fish evolve into amphibains, but what traits do we select to define tetrapod, and also, at what point can one of the nested clades (groups nade up of a common ancestor and all its descendents) be called a tetrapod? This is shown by the following diagram, from a pdf by Dr Stuart Sumida (Biology 680 - Advanced Topics in Evolution - Evolution of the Vertebrate Limbs - left). This whole diagram could be cosnidered a zooming in or higher resolution focus on the drawing of Ichthyostega in the previous diagram. Here A constitutes the actual cladogram itself, which is a hypothesis on phylogenetic relationships, not an actual evolutionary tree (it is easy to get these two ideas confused). Note that this is only one possible cladogram out of many. B is a dendrogram, or chronogram, an evolutionary tree, based on cladogram A, but plotted against geological time. Incidentally one could reverse the position of Colosteidae and Whatcheeriidae, and even make the latter a grade - that would get rid of some of that excessive ghost lineage (the dotted lines not represented by the fossil record). The six drawings on the right are representative early tetrapods, with the most basal (in this case the advanced sarcgopterygian fish Panderichthys) and the most derived or advanced at the bottom. Not only is it difficult to decide at what point a fish becomes a tetrapod, perhaps because the fossils may be incomplete, or there could be disagreement on interpretation, but even where there is agreement on the fossils, there is still a difference of opinion on where tetrapoda starts, according to one's preferred method of phylogenetic nomenclature.
Choice of particular methodology gives certain advantages, but also comes with disadvantages. Evolutionary systematic definitions of amphibia or reptilia may tend to vagueness or ambiguity, but cladistics can be detrimental for the opposite reason: its formalism can be excessively restrictive. Whether having a group of animals or plants (like, say, reptiles, or gymnosperms) rendered invalid because it has descendents that are different to it, is a good or a bad thing, is up to the individual to decide. There is an interesting comment regarding this on the Wikipedia page devoted to phylogenetic nomenclature. The editors there have provided an article that does not take sides or polemically push one position as best (an attitude the present author finds all too prevalent on discussions on phylogeny, evolution, nomenclature etc). They give an analogy with chemistry. To quote:
Proponents of phylogenetic nomenclature claim that, as rank-based nomenclature does not delimit taxa precisely, its definitions will need to change as the science of biology advances. They see their approach as preferable in that their definitions are not susceptible to this kind of instability. It is a disputed matter whether such changes are characteristic of sciences outside of biology and whether, if so, biology should follow the example of such sciences. Chemistry has been used as an example.
Michel Laurin, one of the foremost advocates of phylogenetic nomenclature, considers that the concept of a chemical element has been stable ever since Dmitri Mendeleev put forth the periodic table in 1869. Biology should, on Laurin's view, follow the example of chemistry and define its terms as precisely as possible. (Laurin 2008)
The historian and philosopher Thomas Kuhn argued that changing the meaning of established concepts is central to significant advances in science (Kuhn 1962). Prior to John Dalton's work, he pointed out, the criteria for something's being a chemical compound were such as to include salt water; by new criteria adopted afterwards, this fluid was excluded. (Kuhn 1962)
Michael Benton, a prominent defender of rank-based nomenclature, regards biology as an endeavor very different from chemistry. Chemical classification, as he sees it, circumscribes entities in terms of properties that enter into knowable laws; biology, lacking such laws, must look to the usefulness of classifications. From this perspective, he argues, it is less important that the definition of a term classifying organisms remain constant than it is that the term continue to apply to most of the same organisms. As he views phylogenetic nomenclature as seeking the former kind of stability and rank-based nomenclature as seeking the latter, he considers rank-based nomenclature to be preferable (Benton 2000)
Because of these two different definitions, we can still refer to linnaean-evolutionary taxa and concepts, including similarity, evolution through deep time, supra-specific ranks, and groups that evolve into totally different groups, when the emphasis is on classification applying to particular group of organisms (in this case, Class Amphibia). At the same time, we can also avail ourselves of the statistical and methodological rigour that cladistics provides, when emphasise is on the precise phylogenetic definition of a particular clade (in this case, Clade Tetrapoda). Even though the two systems of classifying life are incompatible, that does not mean that one is wrong and the other right. They can both be right, but in different ways
The problem with relying only on one particular methodology and paradigm can be illustrated with a simple example. In cladistic nomenclature, because of the prohibition against recognition of paraphyletic groups, there is literally no term for the group of transitional forms (Ichthyostega, Acanthostega, etc) intermediate between "fishapod" (very tetrapod-like lobe-finned fish, like Panderichthyes and Tiktilaak) and amphibian-proper (four-legged semi-aquatic animals). Sure, you can say "tetrapod", but that refers to reptiles, birds and mammals as well. The term "basal" is often used by paleo enthusiasts in a vague way to mean the same as "primitive" did under evolutionary systematics. Which is fair enough, but it has nothing to do with the precision of definition that otherwise comes with cladistics. So "basal tetrapod" does not work here either; properly speaking it refers either the node at the bottom of the tetrapod cladogram, or the first terminal to branch off. Normally stem taxon would be the right choice, but because of the ambiguity around the use of the word tetrapod, "stem tetrapod" can mean anything - fishapods, bapheitids, even (in some phylogenetic trees) temnosondyls, .... Because science is about classification, this is a curious, and indeed irritating, lack. That is why there is no simple replacement for Ichthyostegalia (link, link), which can be used to cover all these early transitional forms (not only Acanthostega and Ichthyostega, but also Sinostega, Hynerpeton, and (if it does not belong in a more advanced category) Tulerpeton.
Apart from this, and differing methodological perspectives aside, it doesn't really matter, as Shakespeare informs us, if we refer to primitive amphibians as class Amphibia or basal or stem group tetrapods. In the following pages we will consider more about these ancestors of all land-living animals (as well as of animals that returned to the sea). And here we leave Linnaean ranking in favour of cladistic trees and definitions, beginning with the rather sticky point of the definition of clade Tetrapoda.
MAK111110
Although the earlier classification of subclasses and orders given above no longer can accommodate modern discoveries, with only Temnospondyli (the topic of the next unit) being retained; see Benton 2005 for an updated version.
Quizz results:
If you answered (a) you are a linnanean/evolutionary systematist
If you answered (b) you are a cladist
If you answered(c) you like flying boats
Page by MAK111113