|
|
Temnospondyli |
| The Vertebrates |
Assorted basal temnospondyls |
Temnospondyli: Assorted primitive (basal) temnospondyls
Taxa on this Page
- Balanerpeton X
- Capetus X
- Dendrerpeton X
- Iberospondylus X
- Saharastega X
Assorted basal temnospondyls
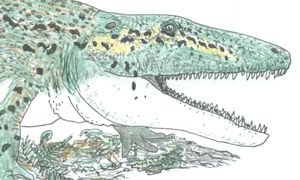
Capetus palustris, life reconstruction, copyright Darren Naish © 2007
From Darren Naish's blog Tetrapod Zoology: Temnospondyls the early years (part II) ( July 9, 2007 ):
Last time we looked at the edopoids, perhaps the most basal temnospondyl clade: here we look at the rest of the basal forms. Scary predators, marine piscivores, late-surviving relics, and some unfortunate beasts burned alive in forest fires...
Studies on temnospondyl phylogeny mostly agree that 'post-edopoid' temnospondyls form a clade, the most basal members of which include Capetus, Dendrerpeton and Balanerpeton (Milner & Sequeira 1994, 1998, Holmes et al. 1998, Ruta et al. 2003a, b) [though some workers have found some of these taxa to be more basal than edopoids (Steyer et al 2006]. In contrast to the condition in edopoids, the interpterygoid vacuities of 'post-edopoids' are rounded at their anterior ends, the jugal (the bone that forms the cheek region) is shortened, and the configuration of skull bones is overall less archaic and fish-like.
Among these basal 'post-edopoids', Dendrerpeton (from the Late Carboniferous of Nova Scotia and Ireland) had a rather large skull with laterally facing orbits, a short body, and well-developed, robust limbs. It lacked lateral line canals and grew to c. 1 m. These features suggest that it was predominantly terrestrial (Holmes et al. 1998). Several species have been named (Milner 1996). Studies of a well-preserved, three-dimensional ear indicate that Dendrerpeton had a frog-like tympanum (ear drum) suited for the perception of airborne sounds (Robinson et al. 2005). This discovery provides support for the idea that lissamphibians descend from temnospondyls (a subject we'll come back to much later), and indicates that at least some basal temnospondyls were listening to noises. But what were they listening to? Were they vocalizing, perhaps during the breeding season? It's previously been argued that the temnospondyl stapes was too massive to support a tympanum (see Laurin & Soler-Gijón 2006), so note that we have to be cautious in inferring the presence of a tympanum in all temnospondyls.
 The Canadian Dendrerpeton specimens come from the famous Joggins Tree Stump Locality where the fossils of several tetrapod taxa (and the earliest land snails) have been discovered inside the hollowed trunks of lycopsid trees. It used to be thought that the animals had fallen into these natural traps and eventually died there of starvation, but the presence of abundant charcoal within the deposits now raises the possibility that the animals were taking refuge from forest fires, and that at least some of these unfortunate creatures were roasted alive (Falcon-Lang 1999, Scott 2001) [the adjacent picture depicts the early reptile Hylonomus, trapped in a hollow tree stump and about to be killed as a forest fire advances overhead].
The Canadian Dendrerpeton specimens come from the famous Joggins Tree Stump Locality where the fossils of several tetrapod taxa (and the earliest land snails) have been discovered inside the hollowed trunks of lycopsid trees. It used to be thought that the animals had fallen into these natural traps and eventually died there of starvation, but the presence of abundant charcoal within the deposits now raises the possibility that the animals were taking refuge from forest fires, and that at least some of these unfortunate creatures were roasted alive (Falcon-Lang 1999, Scott 2001) [the adjacent picture depicts the early reptile Hylonomus, trapped in a hollow tree stump and about to be killed as a forest fire advances overhead].
Another basal temnospondyl is Balanerpeton woodi from the Viséan of East Kirkton in Scotland, a famous locality that has yielded a phenomenal diversity of Carboniferous invertebrates, fishes and early tetrapods. With a length of about 50 cm, Balanerpeton was superficially like a big salamander but it was odd in that, while the 40-42 teeth lining each half of the upper jaw were small, the 25-30 in each half of the lower jaw were much longer. The absence of lateral line canals, ossified wrist and ankle bones, and the apparent presence of eyelids indicate that it also was a terrestrial animal, although its larvae were aquatic (Milner & Sequeira 1994) [Balanerpeton skeletal reconstruction and life restoration immediately below is from Milner & Sequeira (1994), and borrowed from here].
 Also somewhere around the base of Temnospondyli was Capetus palustris from the famous Upper Carboniferous Nýřany desposits of the Czech Republic: this is one of the richest Late Carboniferous fossil sites in the world, yielding at least 700 tetrapod fossils, discovered from the 1870s onwards. Previously, Capetus was regarded by some authors as an edopoid, close either to Edops or to the cochleosaurids, but recent studies have shown that it is not an edopoid, being closer to Balanerpeton (Steyer et al. 2006).
Also somewhere around the base of Temnospondyli was Capetus palustris from the famous Upper Carboniferous Nýřany desposits of the Czech Republic: this is one of the richest Late Carboniferous fossil sites in the world, yielding at least 700 tetrapod fossils, discovered from the 1870s onwards. Previously, Capetus was regarded by some authors as an edopoid, close either to Edops or to the cochleosaurids, but recent studies have shown that it is not an edopoid, being closer to Balanerpeton (Steyer et al. 2006).
Capetus was a fairly scary looking, broad-headed predator with a deep posterior lower jaw: its skull was about 40 cm long, suggesting a total length of c. 1.5 m. As Sequeira & Milner (1993) noted, this makes Capetus one of the largest tetrapods in the Nýřany assemblage. Its teeth were mostly subconical, but those in the anterior part of the lower jaw seem to have been slightly laterally compressed, and possibly with weakly developed keels. As is the case in edopoids and some other basal temnospondyls, its skull bones lack lateral line canals. Sequeira & Milner (1993) suggested that Capetus was an alligator-like amphibious predator specializing on slow-moving tetrapod prey, and that it exploited a different lifestyle from that pursued by its contemporaries, the cochleosaurid Cochleosaurus, and the baphetids Baphetes and Megalocephalus. It was apparently rare in the fauna, with only eight fossils out of 700 from the Nýřany assemblage belonging to this taxon.

Christian Sidor with Saharastega skull. |
Among the most surprising of the basal temnospondyls is the recently described Saharastega moradiensis from Niger. The big deal is that - while Saharastega is apparently way down near the base of Temnospondyli (and hence close to the Carboniferous taxa Balanerpeton and Capetus) - is it from the Upper Permian Moradi Formation, and hence was very much a 'late-survivor', hanging on for long, long after other basal forms had bit the dust. As we saw in the edopoid article, the Moradi Formation seems to contain a strongly provincial, relictual fauna. Saharastega had a fairly nondescript, flattish and subtriangular skull with widely separated and laterally-facing orbits located close to the skull margins. The tabular horn - a pointed projection, growing from the tabular bone at the rear corner of the skull - was particularly odd in this taxon in being directed laterally, rather than posteriorly, and the jaw joint was positioned unusually anteriorly. These peculiarities suggest that Saharastega was doing something interesting, but we don't know what that was. Although originally argued to belong to Edopoidea (Sidor et al. 2005), restudy has shown it to be outside of this clade (Steyer et al 2006).
 Another basal temnospondyl, the Upper Carboniferous Spanish taxon Iberospondylus schultzei, is interesting in that it was discovered in sediments deposited in coastal marine waters (Laurin & Soler-Gijón 2001, 2006). In contrast to the taxa we've looked at so far, it possesses lateral line canals, and therefore was very likely to have been aquatic (though we'll return later to how reliable lateral line canals are in demonstrating aquatic habits). Furthermore, the articulated condition of one of the specimens indicates that little post-mortem transport had occurred, so Iberospondylus was local to the environment in which it was preserved. It's been known for some time that at least some temnospondyls were marine animals (and we'll look at these other marine temnospondyls in a later post), but the basal position of Iberospondylus within temnospondyl phylogeny suggests that members of the group were able to inhabit the marine environment very early on in the group's history. As Laurin & Soler-Gijón (2001) argued, there are indications that this might have been true of even more basal tetrapods: if so, this would explain how Devonian tetrapods became near-globally distributed so early on in their evolution [Iberospondylus skull shown in adjacent image].
Another basal temnospondyl, the Upper Carboniferous Spanish taxon Iberospondylus schultzei, is interesting in that it was discovered in sediments deposited in coastal marine waters (Laurin & Soler-Gijón 2001, 2006). In contrast to the taxa we've looked at so far, it possesses lateral line canals, and therefore was very likely to have been aquatic (though we'll return later to how reliable lateral line canals are in demonstrating aquatic habits). Furthermore, the articulated condition of one of the specimens indicates that little post-mortem transport had occurred, so Iberospondylus was local to the environment in which it was preserved. It's been known for some time that at least some temnospondyls were marine animals (and we'll look at these other marine temnospondyls in a later post), but the basal position of Iberospondylus within temnospondyl phylogeny suggests that members of the group were able to inhabit the marine environment very early on in the group's history. As Laurin & Soler-Gijón (2001) argued, there are indications that this might have been true of even more basal tetrapods: if so, this would explain how Devonian tetrapods became near-globally distributed so early on in their evolution [Iberospondylus skull shown in adjacent image].
This ends our look at the most basal members of Temnospondyli - or, at least, it does according to the phylogenetic schemes I've decided to follow. You will note that at least some of these animals, including the edopoids, Dendrerpeton and Balanerpeton, were apparently terrestrial or mostly terrestrial. It is inferred that a total length of perhaps 40 cm or so was primitive for the group, but a size of 1.5 m or more was evolved within Edopoidea, and also exhibited by broad-skulled Capetus. Some basal temnospondyls were aquatic, and even marine, and some hung on until as late as the Late Permian: about 40 million years longer than we'd thought prior to 2005.
Copyright Darren Naish © 2007, republished with permission
Problems in basal temnospondyl relationships
Although there is agreement on what constitute basal (primitive) and what the more advanced memebers of the temnospondyl family tree - for example both the edopoids and the various genera recviewed here are all recognized as basal - the precise evolutionary relationships among these various basal forms remain controversial. It may be that, instead of a number of well demarcated groups, there is an evolutionary "bush" of many similar forms during the Carboniferous, all of which share a number of primitive features, and perhaps also evolved specialized features in parallel. No doubt future discoveries will clear things up somewhat, but for the present we are faced with a number of conflicting cladograms and evolutionary trees. A few of these are shown below:
For now, we have adopted the sequence of edopoids first (shown on the previous page) and then various assorted forms like Dendrerpeton (this page), and then higher temnospondyls but this arrangement could just as easily been reversed (e.g. Steyer et al. 2006, above). Or as a third option, there may be some very basal forms, then the edopoid clade, then some even more derived but still basal lineages, and finally the "higher" temnospondyls (e.g. Laurin and Steyer 2000 excluding the Limnarchia). And until there's some serious resolution of these problems, we just have to resort to inserting question marks!
MAK090710
Descriptions
Balanerpeton woodi:
Phylogeny: Temnospondyli::: ((:::(Euskelia + Limnarchia)) + *) - See comments on early temnospondyl phylogeny
Characters: Large external nares, large interpterygoid vacuities, ear with large tympanic membrane with rod-like stapes, absence of lateral line sulci, ossified carpals and tarsals (Milner & Sequeira 1994, although Laurin & Soler-Gijón 2006 reject the idea that many temnospondyls possessed a tympanum).
Comments: Named after its discoverer Stanley Wood, Balanerpeton woodi is the most common tetrapod iwhose remains hare found at the East Kirkton locality. It appears similar to Dendrerpeton. The structure of the ear suggests the ability to hear high-frequency sound. This was clearly a terrestrial animal; not only is the locality a terrestrial one (terrestrial arthropods but no fish are known from East Kirkton), but Balanerpeton lacked lateral line sense organs or ossified branchial system, the bony (rather than cartiligenous) wrists and ankles, and seems to have possessed eyelids.
References: Milner & Sequeira 1994
Links: Tetrapoda - Balanerpeton, Wikipedia
MAK090723
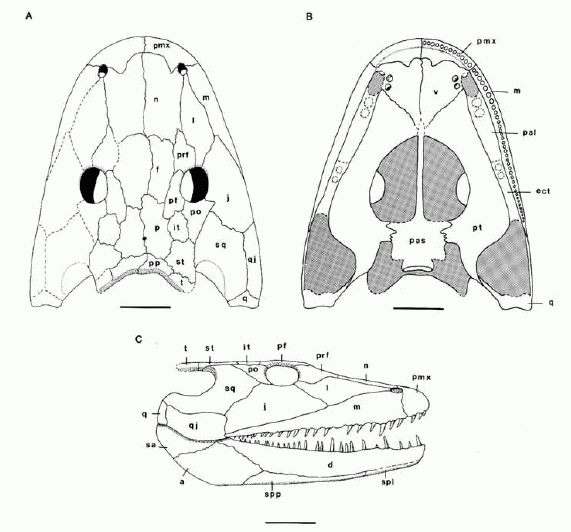
Capetus palustris, reconstruction of skull of large specimen in dorsal (A), palatal (B), and lateral (C) views. Scale bar is 5 cm. Sequeira & Milner1993 p.672 |
Capetus palustris:
Late Carboniferous (Westphalian D - Late Moscovian) of Nýřany, Czech Republic
Phylogeny: Temnospondyli::: ((:::(Euskelia + Limnarchia)) + *) - See comments on early temnospondyl phylogeny
Size. Skull length of upto 40 cm. Sequeira & Milner 1993 estimate from there a total length of 1.5 meters, although if we assume Balanerpeton or Dendrerpeton-like proportions, the length was probably a little larger.
Comments: A generalized basal temnospondyl, it has no features that would place it either with the Edopoidea or with the Eryopoidea. Instead it would seem to be a transitional form more advanced than the former but not as specialized as the latter. Shared its environment with another large temnospondyl, Cochleosaurus. It was among the largest animals of its environment, and probably filled an Eryops-like role of amphibious apex preditor.
Reference: Sequeira & Milner 1993
MAK090724
 Dendrerpeton:
Dendrerpeton:
Range: Early? to Late Carboniferous of North America & Ireland(?)
Phylogeny: Temnospondyli::: ((:::(Euskelia + Limnarchia)) + *) - See comments on early temnospondyl phylogeny
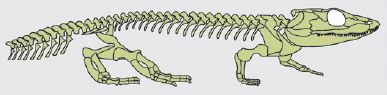
Skeleton of Iberospondylus Dendrerpeton, from diagram in Schoch 2008 p.221
|
Characters: Up to 100 cm; large laterally facing orbits; $ jugal narrowing to a point, making a point contact with the lacrimal; large, rounded "otic notch" in squamosal; stapes massive [suggesting support structure, not related to hearing -- but the "massive stapes" may be controversial, see Biology 356]; possible lanceolate expansion on anterior tip of cultriform process; lateral line sulci absent; short presacral column of 24 vertebrae, less than twice skull length; large stout limbs.
Note: according to one source, these characteristics "suggest a terrestrial lifestyle distinct from the aquatic and semiaquatic adaptations of most contemporary Carboniferous amphibians"
Image: photograph colorized and labelled per the diagram at Dendrerpeton and Joggins, Nova Scotia.
Links: Dendrerpeton and Joggins, Nova Scotia; Joggins Cliffs; Biology 356; Phylogeny and Apomorphies of Temnospondyls; Fundy Geological Museum, Paleontology, Fossils, Vacation ...; Localities of the Carboniferous-; Ontogeny and phylogeny in temnospondyls- a new method of analysis pdf of Steyer, 2000). ATW031216.
Iberospondylus schultzei:
Late Carboniferous (Stephanian C - Early Gzhelian) of Spain (Puertollano Basin.)
Phylogeny: Temnospondyli::: Euskelia + * - synapomorphies indicate that Iberospondylus is more closely related to eryopoids and dissorophoids than to edopoids, although the phylogeny, like others on temnospondyls, is not robust (Laurin & Soler-Gijón 2006). See also comments on early temnospondyl phylogeny
Comments: Previously all Permo-Carboniferous amphibians were thought to have been fresh-water only, but Iberospondylus apparently lived in a coastal marine environment. Spanish The first and to date only known occurrence of a Paleozoic amphibian in Spain.
References: Laurin & Soler-Gijón 2001, Laurin & Soler-Gijón 2006 - abstract.
MAK090723
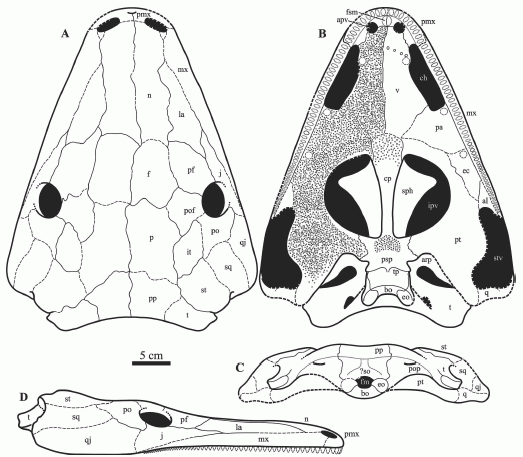
Reconstruction of the holotype skull of Saharastega moradiensis in dorsal (A), palatal (B), occipital (C), and lateral (D) views. There is little or no evidence for the presence of palatal fangs, although they are tentatively included in the reconstruction. Scale bar is 5 cm. Damiani et al 2006 p.564
|
Saharastega :
Phylogeny: Here there are two options to choose from:
Damiani et al 2006: Temnospondyli:: (Capetus + (Euskelia + Limnarchia)) + (Edopoidea + *)
i.e. Saharastega and Edopoids form a clade which is the sister group to all other Temnospondyls; in other words the most primitive or basal outshoot of the temnospondyl ancestry
or Steyer et al. 2006 : Steyer et al. 2006: Temnospondyli:: (Edopoidea + Eryops) + * Saharastega represents a primitive but not the most primitive form; Edopoids + Saharastega are a grade not a clade, and Saharastega is the sister group to the Edopoids and Eryopoids.
Characters: Nostrils narrow and elongated; premaxillae-maxillae articulation via extensive tongue-and-groove contact; exceptionally large tabular "horns" directed both laterally and ventrally; dorsal region of occiput comprises a complex "occipital plate" . (Damiani et al 2006, p.561)
Comments: A large but very primitive temnosopondyl; a Carboniferous-grade "living fossil" that survived till the end of the Permian, thanks to geographical isolation. More terrestrially adapted than its equally primitive crocodile-like cousin Nigerpeton (Steyer et al. 2006 p.226). Curiosuly, there seems to have only been the small teeth around the margins of the mouth, with no trace of vomerine and palatine tusks that are always present in temnospondyls (Damiani et al 2006 p.567). This may be a result of unfavourable preservation. Or it may be that Saharastega was simply specialized as a harmless fish-eater (although in that case why the terrestrially adaptations?)
Reference: Damiani et al 2006, Steyer et al. 2006, Sidor et al. 2005
MAK090710

