
| Temnospondyli | ||
| The Vertebrates | Metaposauroidea |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Tetrapoda
├─┬─Lepospondyli
│ └─Reptiliomorpha
│
└─Temnospondyli
├─Edopoidea
└─┬─Dvinosauria
└─┬─Euskelia
│ ╘═Dissorophoidea
│ └─Lissamphibia
└─Stereospondyli
├─Rhinesuchidae
└─┬─Lydekkerinidae
├─┬─Plagiosauroidea
│ └─┬─Rhytidosteidae
│ └─Brachyopoidea
└─┬─Capitosauria
└─Trematosauria
├─Trematosauroidea
└─┬─Metoposauroidea
├─Inflectosaurus
└─┬─Latiscopidae
└─Metoposauridae
├─Apachesaurus
├─Arganasaurus
├─Dunuitosaurus
├─Koskinonodon
└─Metoposaurus
|
Contents
Index |
Imagine an animal sitting at the bottom of a Triassic swampy pond or shallow lake, vaguely resembling a huge and quite flat frog, except with small hind legs and stubby tail; the whole creature some 2 meters in length. The head is almost pancake flat, the small upward facing eyes far forward and close to the snout. The scaly skin is a green or brown or mottled color to blend in with the murk, mud, and weeds among which the creature spends most of its time, completely motionless. Now and then it moves is to push off the bottom with a kick of a hind leg and a brief tail wriggle, take a gulp of air from the surface, and resume its position. Occasionally an unwary fish, or maybe a large crustacean, small amphibian, or juvenile phytosaur will swim by too close, wherupon the huge trapdoor mouth snaps open and shut and the luckless prey is gone.
If you can imagine that, you can imagine a metoposaur, or at least what I would think they might look and act like. Metoposaurs were one of the success stories of the Carnian (early part of the late Triassic) age, appearing out of nowhere to attain almost worldwide distribution (only absent in South America (South-West Pangea), perhaps because of geographical obstacles). Then, after some ten million years of success, they markedly declined, victim of the end Carnian extinction event. A few diminutive forms contuinued into the following, Norian , age, including a quite successful terrestrial type, but they too dissapeared, leaving no descendents of this short-lived but successful line.
What is very interesting about the Metoposaurs is that they weren't the only giant, flat-bodied, short-tailed, pancake-headed, bottom dwelling amphibians making life miserable for the smaller denizens of Triassic lakes, rivers and swamps. Move the eyes further towards the rear of the skull, make the body not quite as flat, you have a Capitosaur. Or, make the head very short, but still wide, so it is several times wider than long, and keep the body flat, add big feathery external gills, the result is a Plagiosaur. Why was it that three totally independent lines of temnospondyl each convergently evolved the same size and form, and how were they able to co-exist without out-competing each other?
Clearly Carnian ecosystems were rich, complex, and highly diverse; indeed the Cranian in many ways was the apex of the Triassic. I am reminded of the mighty Carboniferous coal swamps, where there were always a number of quite distinct but convergent large pike- or eel-like fish and amphibians (Xenacanthida, Rhizodontiformes, Crassigyrinidae, Colosteidae, Eogyrinidae, etc), each co-existing quite happily within specific niches, even though they would seem to follow the same lifestyle. But although both biomes show the same example of convergence among multiple aquatic clades, there the parallels ended. In the Carboniferous, a narrow, seperentine-like body was necessary to navigate the submerged logs and weeds to actively search for prey; in the Triassic, it seems like a bottom-dwelling flat-bodied ambush-predator was the way to go.
The end of the Triassic didn't mean the end of the amphibian giants. One clade, the brachyopoids continued right until the Cretaceous, and in at least one case attaining gigantic size. But they were never common or widespread except perhaps locally; the post-Triassic world was a very different place, and the time in which large amphibians dominated aquatic environments was gone forever.
MAK090808
Metoposauroidea: Almasaurus, Koskinonodon. Definition: Almasaurus + Metoposaurus. Yates & Warren (2000).
Range: Middle Triassic (Ladinian) to Late Triassic (Rhaetian). Probably cosmopolitan.
Phylogeny: Trematosauria:: (Plagiosauroidea + Rhytidosteidae + Brachyopoidea))) + *: Inflectosaurus + (Latiscopidae + Metoposauridae)
$ infra-orbital sulcus with a step-like flexure between orbit & naris [this appears to mean that the sensory line canal passing below the orbit makes a sharp medial or dorsal turn just anterior to the orbit, then an opposite turn anteriorly, to pass just outside each naris. However, this interpretation may be erroneous, since Batrachosuchus (=Batrachosaurus) Plagiosauroidea) and even Neldasaurus (Dvinosauria) seem to have the same feature]; $ length of posterior skull table >90% of width; $ "gutter" bordered by ridges at margin of otic notch; exoccipital-pterygoid suture visible in ventral view; paired anterior palatal fossae; $ keeled lateral margin of clavicle.
Yates & Warren (2000) [comparative remarks based on figures in Carroll 1988) from Watson (1956) and from Chase (1965), vide Carroll 1988)].
(ATW 000213)
Inflectosaurus: Shishkin 1960. S. amplus Shishkin 1960.
Range: Early Triassic of Russia, Yarenskian Gorizont (Astrkhan)
Phylogeny: Metoposauroidea: (Latiscopidae + Metoposauridae) + * . [S02]
Characters: skull up to 70 cm [S+00]; orbits small [S+00]; pineal foramen close to middle of orbit - occiput distance [S+00]; quadrates posterior to occipital condyles [S+00]; preotic pterygoid ascending ramus strongly developed [S+00].
References: Shishkin et al. 2000) [S+00]; Steyer (2002) [S02]. ATW020721.
Latiscopidae [Almasauridae]: Latiscopus disjunctus, Almasaurus habbazi.
Range: Late Triassic (Late Carnian to Early Norian). SW USA (Latiscopus) and Morrocco (Almasaurus is known from the Lower Irohalene Member - Early Late Carnian of Morocco)
Phylogeny: Metoposauroidea: Inflectosaurus + (Metoposauridae + *)
Characters: slightly elongate and narrowly pointed muzzle,small lateral orbits, prominent dermosensory canals
Size: overall length about a meter
Comments: aquatic piscivores
References: Milner 1994 p.10,
MAK090724
Metoposauridae: Watson, 1919 Metoposaurus, Koskinonodon.
Range: Late Triassic (Carnian to Norian). Probably cosmopolitan.
Phylogeny: Metoposauroidea: Inflectosaurus + (Latiscopidae) + * : Dunuitosaurus + Metoposaurus + Koskinonodon
Characteristics: Lachrymal separated from the orbital margin by broad prefrontal-jugal suture; small widely spaced nares; continuous lateral-line loop behind the orbit [Milner 1994]
Introduction: A group of large flat-headed aquatic tetrapods. Although in appearance very like the capitosaurids in size and body proportions, the metoposaurs belong to a distinct lineage of temnospondyls. The most noticeable distinction (apart from various details of the skeleton) are the more forward position of the eyes. The metoposaurs were large mostly aquatic predators that fed on fish and small animals. As with many Triassic temnospondyls tetrapods the head was large and flat, with the eyes looking upwards. The creature probably spent a great deal of time submerged and motionless, waiting for an unwary fish or smaller tetrapod to swim past. The jaw was lined with teeth, and there were large teeth on the palate. Metoposaurs were strong swimmers, but would have been very clumsy on land, and it is likely that they ventured from water rarely, if at all.
Large numbers of fossil specimens have been found crowded together, dying when the ponds and lakes in which they lived dried up, and preserved when the mud that covered them hardened into rock. (MAK 980114)
Comments: These large successful aquatic animals are entirely limited in range to the Late Triassic, and were most common durintg the Carnian, when they achieved almost cosmopolitan distribution (they are however unknown from South America (SW Pangea) where their ecological role was taken by mastodonsaurs).
Metoposaurs were large amphibians; averaging about 2 meters in length, the largest may have reached 3 meters, rivalling the larger Capitosaurs in size. Koskinonodon perfectum (formerly known as Buettneria) had a skull 65 cm long and would have no doubt been able to hold its own against all but the largest phytosaurs. At the other end of the scale, the small, elongate, terrestrial Apachesaurus, with a skull less than 20 cm in length, seemed to have been survived after the larger aquatic types died out.
Relationships between metoposaur species
Attempts at working out the phylogeny of these animals remain controversial; as with temnosopondyls in general, there seems to be little agreement regarding the relationship between the different Metoposaur taxa, although the work of Sulej 2002 and 2007 has helped clarify the status of Metoposaurus. The following trees are presnted for comparison:
|
■──┬─Dunuitosaurus
└─┬─Apachesaurus
└─┬─┬─B.bakeri
│ └─M. diagnosticus
├─Arganasaurus
└─┬─B. perfecta
├─Anaschisma
└─Dictyocephalus
|
■──┬─B. howardensis
├─┬─B. perfecta
│ └─┬─Dunuitosaurus
│ └─┬─Arganasaurus
│ └─Apachesaurus
└─┬─B. bakeri
└─┬─Metoposaurus maleriensis
└─┬─M. d. diagnosticus
└─M. d. krasiejowensis
|
||
Hunt 1989. |
Sulej 2007 ( converted to dendogram). |
MAK090724
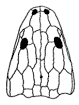 Apachesaurus: Apachesaurus gregorii Hunt, 1993
Apachesaurus: Apachesaurus gregorii Hunt, 1993
Range: Late Triassic (Latest Carnian to Middle Norian). SW N Am. ( Lower and Upper Chinle and Redonda Formations of Arizona and New Mexico, Dockum Group of Texas). This animal is rare in the Carnian, were larger forms dominate, but much more common during the Norian [Milner 1994 p.11]
Phylogeny: Metoposauridae::: *
(see comments on phylogeny)
Synonyms: Kalamoiketor pinkleyi Branson & Mehl, 1929, Anachisma sp., perhaps Dicytocephalus.
reduction of otic notch and body size (Sulej 2007 p.124), Otic notch shallow and rounded [Hunt 1989], elongation of body, vertebral intercentra elongate with excavated articular faces, absense of pleurocentra, skull less than 25 cm [Milner 1994]
Ecological Niche: Terrestrial/Semi-aquatic insectivore/small carnivore
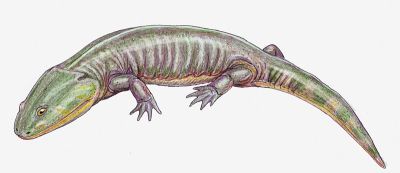 Life reconstruction of Apachesaurus gregorii, by Dmitry Bogdanov (Wikipedia) |
Notes: Apachesaurus is an unusual dwarf metoposaur (skull lengths of less than 20 centimeters) with a number of adaptations for a terrestrial existence (e.g. advanced pelvis). It seems that following the aridification of west equatorial Pangea, this little animal, which was pre-adapted to be less dependent on water than its larger cousins, not only survived but flourished. It is known from abundant intercentra, teeth, and other remains, as well as several partial skulls and skeletons, and other postcrania. Although Lucas 1998 mentions only a single species, and indicates this animal seems to have continued right up until the very end if the Triassic, it is not unlikely that over this extended period of time there were a number of closely related and similar species existing. A more realistic range would be latest Carnian (Adamanian faunal stage) to Early or Mid Norian
Link: Petrified Forest National Park - Apachesaurus (useful reference but much shorter than their page on Koskinonodon); Wikipedia (includes references and useful life reconstruction (left), but nothing much else when I last looked)
MAK090724
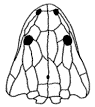
Arganasaurus lyazidi (Dutuit, 1976).
Range: Late Triassic (Upper Irohalene Member - Latest Carnian). Morocco.
Phylogeny: Metoposauridae::: *
(see comments on phylogeny)
reduction of otic notch and body size (Sulej 2007 p.124)
Notes: "Metoposaurus Grade 2" of Milner 1994. Dunuitosaurus ouazzoui (Dutuit, 1976) is included in this clade by of Milner 1994. Hunt 1989 considers that because of primitive features (presence of pleurocentra elements, long intercentra) that species is much more basal. Arganasaurus is distinctively smaller, and has a shallower otic notch, than Dunuitosaurus (Sulej 2007 p.123).
MAK090724
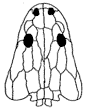

Dunuitosaurus: Dunuitosaurus ouazzoui (Dutuit, 1976) .
Range: Late Triassic (Lower Irohalene Member - Early Late Carnian (Otischalkian of Lucas 1998) of Morocco.
Phylogeny: Metoposauridae::: *
(see comments on phylogeny)
Pleurocentra elements, intercentra long [Hunt 1989]. Tail long (Lambert et al 2001, p.60)
Note: A fairly early form, characterised by primitive features, such as long intercentra. Other Metoposaurs have short intercentra. The long tail implies that Dunuitosaurus was more of an active hunter than a lurk and wait ambush predator.
MAK090808
"Eupelor" bakeri (Case, 1931)
Range: Late Triassic (Earliest Late Carnian - Otischalkian Age) N Am. (Dockum Group, Texas; Fundy Basin, Newark Supergroup, Nova Scotia). - note, the generic designation Eupelor is a provisional one, assuming that this species is not necessarily closely related to K. perfecta.
Phylogeny: Metoposauridae::: *
(see comments on phylogeny)
Synonyms: Buettneria bakeri Case, 1931, Eupelor fraasi jonesi (Case, 1922), Metoposaurus fraasi jonesi (Case, 1922), and Metoposaurus bakeri (Case, 1931) Hunt, 1993], Koskinonodon bakeri
Characters: Anterior shift of lachrymal (Sulej 2007); lachrymal not close to orbit [Hunt 1989], incomplete sensory canal system [Murray 1986], elongate pits in the frontal and postorbital regions, extension of the palatal vacuities anterior to the orbits, narrower and more elongate skull, smaller adult size.
Notes: This species may be more primitive than other Metoposaurs, and it has an earlier stratigraphic range (Sulej 2007 p.123). . In a number of phylogenies it is not necessarily the sister taxon of Koskinonodon perfecta. Murray 1986 suggests that the elongate palatial vacuities and narrowness and small size of the skull indicate a juvenile rather than a distinct species, but other references acknowledge it as a valid species . Juvenile features could simply be a primitive neotonous or paedomorphic form. Also the incomplete lateral-line system is an advanced feature, like the elongate lachrymal entering the orbit, this feature seems to have evolved several times among metoposaurs.
According to Sulej 2007 p.126, Buettneria bakeri initiated a new lineage, in which the lacrimal out of the orbit margin. This was charcterised by a tendency to decreasing size and depth of the otic notch, and elongation of the interclavicle, and culminated in Apachesaurus.
B. bakeri also has a similar sculpture of the centrum of interclavicle to B. perfecta, which has lacrymal forms the margin of the orbit. In both these species the large area of the centrum is charcterised by polygonal pits. This feature distinguishes "Buettneria", from North America from the European and Indian species of the genus Metoposaurus Sulej 2002 p.125
MAK090724
Koskinonodon perfecta Case, 1922
Range: Late Triassic (Late Carnian) N Am. (Chinle Formation, Arizona and New Mexico; Dockum Group, Texas and New Mexico; Popo Agie Formation, Wyoming)
Phylogeny: Metoposauridae::: *
(see comments on phylogeny)
Synonyms: Buettneria Case, 1922 (Mueller, 2007) The generic name being preoccupied, it was replaced with Koskinonodon Branson and Mehl, 1929. Species synonymns: Borborophaqus wyomingensis Branson & Mehl, 1929, Buettneria calgariensis Green, 1954, Buettneria howardensis Sawin, 1945, Buettneria major Branson and Mehl, 1929, Buettneria perfecta Case, 1922, Eupelor fraasi jonesi Colbert and Imbrie, 1956, Eupelor fraasi fraasi Colbert and Imbrie, 1956, Eupelor browni Colbert and Imbrie, 1956, Koskinonodon princeps Branson & Mehl, 1929, Metoposaurus browni Roy Chowdhury, 1965, Metoposaurus jonesi Case, 1920, Metoposaurus fraasi jonesi Roy Chowdhury, 1965, Metoposaurus fraasi fraasi Roy Chowdhury, 1965
 Skeleton of Koskinonodon (Buettneria), Dinosaur Exhibit at the American Museum of Natural History. (Wikipedia) What's wrong with this mount? That's right, the tail is too long! When skeletons of this species were first found, the tail was incomplete. Not knowing how much tail to add, and because the long, serpentine tail looks sexier, it was given a big tail at least in some reconstructions. Compare also the overlong tail of early reconstructuction of Tyrannosaurus rex. In life Koskinonodon almost certainly had a shorter tail, like the closely related Metoposaurus (see sketch of Metoposaurus skeleton). |
Characters: Elongation of lachrymal, shortening of prefrontal, reduction of interclavicle, centre (Sulej 2007), lachrymal entering the margin of the orbit. These featrures are all continued in Metoposaurus (Sulej 2002). The implication being that this form is either ancestral to Metoposaurus (Sulej 2007 p.127) or evolved Metoposaurus-like features in parallel.
Size: average skull length of about 65 centimeters. Assuming the same proportions as Metoposaurus diagnosticus (see illustration) that gives an overall length of about 3 meters, making it among the largest of the metoposaurs.
Comments: A large and common animal, better known as Buettneria, The weak axial skeleton and intact lateral line system of the skull indicates that Koskinonodon was a fully aquatic amphibian. The teeth are sharp and conical, indicating a diet of fish and other small aquatic animals. The upward facing eyes indicaye an ambush predator, lying in wait in the soft mud at the water bottom and attacking suddenly from below. (NPS website)
Sulej 2007 considers B. howardensis a distinct species of Buettneria, and places this species at the bottom of his non-cladistic phylogenetic diagram (Sulej 2007 p.124). Anon (unsigned NPS website) identifies it with Koskinonodon perfecta
Link: Petrified Forest National Park - Koskinonodon (a detailed page and useful reference)
MAK090724
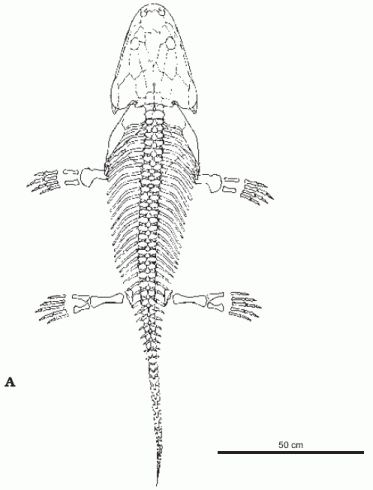  Metoposaurus diagnosticus krasiejowensis. Dorsal (A) and Lateral (B) view of skeleton.. Overall length 1.75 meters. Illustration from Sulej 2007 p.122 |
Metoposaurus Lydekker, 1890
Range: Late Triassic (Late Carnian to Early Norian). Europe and India.
Phylogeny: Metoposauridae::: * : Metoposaurus maleriensis + Metoposaurus diagnosticus
(see comments on phylogeny)
Characters: Lachrymal enters orbital margin as with Koskinonodon Sulej 2002 p.539; interclavicle with relatively long posterior part; small centre consisting of isometric pits; the glenoid of scapula directed posterolaterally; the braincase weakly ossified; the humerus, scapula, and fibula relatively slender. Sulej 2007 p.128
Comments: The German Metoposaurus has traditionally been distinguished from the American Buettneria (now Koskinonodon) on a certain detail of the bones around the eye socket. In Koskinonodon the lachrymal enters orbital margin (this being a specialized feature), whereas (Fraas 1889, Romer 1947) it was always thought that with Metoposaurus this was not the case. Metoposaurus therefore was given a more basal position in all phylogenies, for example Hunt 1989. But in a restudy of the European material, as well as of specimens from Krasiejów in Poland, Sulej 2002 has shown that in all European metoposaurs the lacrymal does actually enters the orbital margin, and in this regard these specimens do not differ from those of Buettneria perfecta (Koskinonodon) from North America. Sulej suggests that the metoposaur skulls studied by Fraas were poorly preserved and so he may have based his interpretation of the shape of the Metoposaur lacrimal largely on the much better preserved Cyclotosaurus fossils, to which he compared it.
In contrast to the lacrimal topography of Metoposaurus and Koskinonodon, in Buettneria bakeri (here "Eupelor"), Apachesaurus gregorii, Dutuitosaurus ouazzoui, and Arganasaurus lyazidi the lacrimal is excluded from the orbital margin. (Sulej 2002 p.539)
MAK090724
Metoposaurus maleriensis: Roy Chowdhury, 1965
Synonyms: Buettneria maleriensis (Roy Chowdhury, 1965) Sengupta, 2002
Range: Late Triassic (Maleri Formation - Early Late Carnian (Otischalkian ) of India. (Lucas 1998 p.366)
Phylogeny: Metoposaurus : Metoposaurus diagnosticus + *
Comments: Identified with Koskinonodon perfecta by Anon (unsigned NPS website), referred to Metoposaurus by Sulej 2007, who presents this species as more primitive than Metoposaurus diagnosticus
MAK090724
Metoposaurus diagnosticus (Meyer, 1842) Lydekker, 1890
Synonyms: Synonyms to this species include Metopias diagnosticus Meyer, 1842; M. stuttgartiensis Fraas, 1913 (nomen dubia) ; M. heimi Kuhn, 1932; and Buettneria perfecta Case, 1922 in part].Range: Late Triassic (Late Carnian to Early Norian). Europe.
Phylogeny: Metoposaurus : Metoposaurus maleriensis + * : M. d. diagnosticus + M. d. krasiejowensis
Characters: Clavicles have long common margin anterior to the interclavicle [Hunt 1989] (only metoposaur with clavicles united in this way); Lachrymal enters orbit (Differs from Apachesaurus; Arganasaurus; Dutuitosaurus; and Koskinonodon bakeri in the position of the lacrimal which enters the orbital margin); area of rounded pits in middle of interclavicle small (Differs from Koskinonodon and Apachesaurus in the smaller area with sculpture formed by the isometric pits of the interclavicle.) (Sulej 2002)
Notes: A common European genus, which is distinct from the American and Gondwanan forms. It is a monotypic genus with a single species Metoposaurus diagnosticus with two subspecies. This animal flourished across a wide area during the Carnian, before dying out at the end of the age.
Reference: Sulej 2002
Comment: The type of Metoposaurus stuttgartiensis, from the early Norian Lehrbergstuff of Stuttgart-Sonnenberg, Germany, is the only known Norian metaposaur from Europe. Although too fragmentary to be diagnostic, I wouldn't be surprised if this turned out to be a distinct species. Like Apachesaurus, it was a relatively small animal [Milner 1994]
MAK090724
Metoposaurus diagnosticus diagnosticus (Meyer, 1842)
Range: Late Triassic (Early Late Carnian). Western Europe.
Phylogeny: Metoposaurus diagnosticus + * : M. d. krasiejowensis + *
Notes: the western subspecies
Reference: Sulej 2002
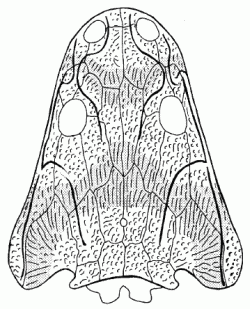 Metoposaurus diagnosticus krasiejowensis. Zones of intense growth shaded. Illustration from Sulej 2002 p.545 |
Metoposaurus diagnosticus krasiejowensis. Sulej, 2002
Range: Late Triassic (Otischalkian (Lucas 1998) /Early Ischigualastian (Langer 2005) / Early Late Carnian - Drawno Beds - Silesia in Poland). Eastern Europe
Phylogeny: Metoposaurus diagnosticus : M. d. diagnosticus + *
Characters: Differs from Metoposaurus diagnosticus diagnosticus in the much shorter pre-pineal part of parietal, and the larger expansion angle of sutures separating the parietal from the supratemporal (Sulej 2002 p.545)
Notes: the eastern subspecies, a more advanced species than M. d. diagnosticus The age of the Krasiejów spinicaudatan assemblage probably lies within the middle-late Carnian, which would mean that the two subspecies of M. diagnosticus are not chronosubspecies but geographic races. (Sulej 2007 pp.127-8).
Reference: Sulej 2002, Sulej 2007
MAK090724
checked ATW040722, last modified MAK090808
{kind=link}
{kind=link}