Vertebrata
|
Abbreviated Dendrogram
|
Contents
|
Taxa on This Page
- Vertebrata
 |
Representative vertebrates. Clockwise, starting from top left: 1. Fire Salamander (Salamandra salamandra) 2. Saltwater Crocodile (Crocodylus porosus) 3. Southern Cassowary (Casuarius casuarius) 4. Black-and-rufus Giant Elephant Shrew (Rhynchocyon petersi) 5. Ocean Sunfish (Mola mola).
|
The most basal group of vertebrates are the Hyperoartia lampreys) which survive as disgusting, but rather interesting, blood-sucking parasites on fish. In addition, the vertebrates produced the Euconodonta, the Pteraspidomorphi, and a vague, perhaps polyphyletic group referred to for the time being as "Thelodonti". These will be discussed in more detail in other sections.
Work on what is now the vertebrates section of Palaeos began less than five years ago. At that time, the consensus view of early vertebrate evolution was not far from the cladogram reflected when the predecessor to this site first went on line. The standard texts generally reflected craniates evolving sometime in the Ordovician, with vertebrates arising somewhat thereafter, perhaps during an apparent secondary "explosion" in the Silurian. Eventually, the Osteostraci emerged, and some Osteostracan or a closely related forms developed jaws and paired pectoral limbs -- all at about the same time -- resulting in another quick radiation of gnathostome forms during, perhaps, the Late Silurian. At that time, conodonts were not considered chordates -- much less vertebrates -- and the thelodonts were considered as little as possible, since they made no phylogenetic sense. For that matter, the thelodonts still make very little sense.
Nonetheless, things have changed a good deal, and -- although there is probably not yet a new consensus -- it has been necessary to change the very backbone of the vertebrate cladogram (to introduce a serious redundancy). After a recent series of significant revisions, the story unfolds at a much more leisurely pace.
The origin of chordates now seems to date back to Pre-Cambrian times, perhaps
600 My. Haikouella was described by from the Early Cambrian (about 530 My) of South China in 1999. It is a derived chordate, just short of being a crown group craniate. Myllokunmingia, from about the same time and location, probably is a crown group craniate. Shu et al. (1999). Crown group vertebrates, i.e., the last common ancestor of lampreys and gnathostomes, can scarcely have been much later. In fact Haikouichthys, also described in Shu's paper, may well be a very primitive hyperoartian lamprey cousin).
By Furongian times, at 500 My, vertebrates had evolved a variety of forms, including the extraordinarily strange and successful euconodonts. Conodonts have been known for almost as long as there has been paleontology, but their peculiar dentition was all that was known of the group. Since conodont remains are very common, they were much-studied as stratigraphic markers but otherwise little understood. As Donoghue et al. (2000) state in their recent study: "Just a year before the first conodont fossil with preserved soft tissues was found, Müller (1981) compiled a list of groups to which conodonts had been attributed; his list includes at least three kingdoms and almost every major animal phylum." Id. at 192.
It now seems fairly well accepted that conodonts are chordates. The Donoghue group's own detailed cladistic study places them within the vertebrates, just above the Hyperoartia. Interestingly, the authors state that the earliest (Pre-Cambrian to Early Cambrian) conodonts, known as the Protoconodonta, are probably unrelated. They assert that the Early to Late Cambrian Paraconodontida are related, but do not include them in their analysis because there are no known soft tissue remains. Thus their analysis is based only on the relatively derived Euconodonta. If paraconodont soft tissue remains are ever found, the picture may be different.
The exact placement of this group is of some interest because the inclusion of conodonts has other strange effects on the vertebrate cladogram. Donoghue et al.'s final result looks like this:
Note that the headshield forms (osteostracans and their close relatives) are now monophyletic -- no longer gnathostome ancestors. This is odd, but appealing. The Osteostraci have never been satisfactory gnathostome ancestors because they are very highly derived forms. The general direction of osteostracan evolution seems to have been toward a loss of fins and an ever-more benthic existence. Only the most basal forms, like Ateleaspis, really suggest a gnathostome relationship. The separation of anaspids from lampreys is less congenial. Janvier 1996) shows a rather neat series between the anaspids and lampreys which can be seen below. However, this separation does seem to be required by the current data.
Somewhat similar results were obtained by Wilson & Caldwell (1998) in their study of the "fork-tailed" thelodonts, the Furcacaudiformes. The thelodonts, like conodonts, are another old group of misfits. They come in a great variety of forms: some resembling anaspids, osteostracans, and gnathostomes. Thelodonts appear to be united only by lacking large dermal plates and having very small scales somewhat similar to those of primitive sharks, and have been widely believed to be para- or polyphyletic. See, e.g. Thelodonti. Some appear to have paired fins (located over the gill slits) and other advanced features, including a well-developed anal fin and possible fin spines. Unfortunately, since thelodonts lack large dermal plates or massive cartilaginous headshields, they are rarely found as articulated specimens. Wilson & Caldwell, however, used their new data on the aberrant furcacaudiforms to try to anchor the group. Although some of their conclusions are too difficult to accept at the moment, they, like Donoghue et al., also conclude that Loganellia and other thelodonts (e.g. Turinia) are close to the gnathostome base and that that the osteostracans are several steps removed.
Given these results, it may now be possible to advance seriously the hypothesis that the thelodonts are the main stem of vertebrate evolution, and that the other groups are only branches. Consequently, we have taken the liberty of temporarily promoting the thelodonts to "Thelodonti" to include anaspids, gnathostomes, furcacaudiforms, and the headshield forms. This solution is only slightly radical and finds comfortable homes for both of the perennial orphans of early vertebrate paleontology: the conodonts and thelodonts. No doubt these are only foster homes, and some new and different arrangement will be necessary in a few more years, particularly for the conodonts. However this hypothesis surely makes more sense than relegating to footnotes the conodonts, who make up 65% of all non-gnathostome chordate species, and thelodonts, our closest known relatives among jawless fish.
Ed. note - this was written in 2001 - MAK111022.
Update: While vertebrate systematics, like science in general, is likely always to remain in a state of flux, a consensus is emerging regarding the relationships of some of the primitive jawless forms (e.g. Janvier 2007a, Janvier 2007b, Sansom et al 2010). The following dendrogram is from Janvier 2008 fig 1, Sansom et al 2010's tree differs in some respects.
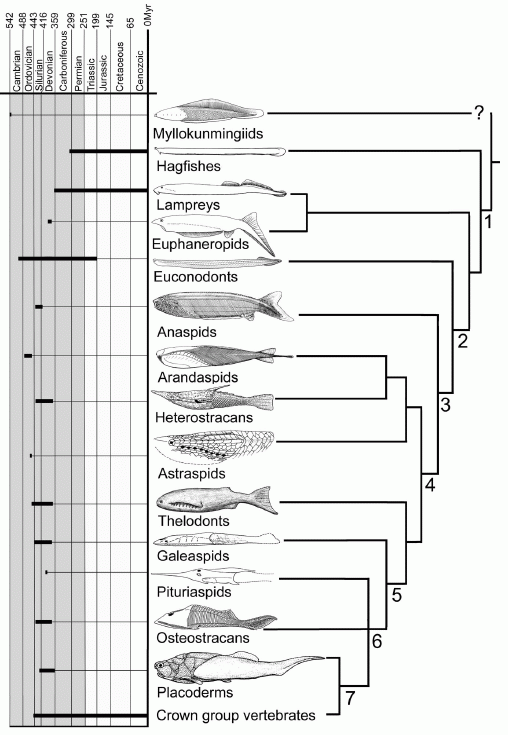 |
Dendrogram showing the distribution of the taxa through time. Stratigraphic range is indicated by bold lines in the time scale to the left, phylogeny by the tree on the right. Major synapomorphies as follows: 1, neural crests, epidermal placodes, fin radials; 2, dermoskeleton in mouth and pharynx; 3, extensive dermoskeleton over the entire body; 4, extensive lateral-line system enclosed in grooves and canals, vertical semicircular canals forming loops, cerebellum; 5, endoskeleton lined with calcified cartilage or perichondral bone; 6, pectoral fins in postbranchial position; 7, jaws.
|
The main difference with earlier phylogenies is that primitive forms like Euphanerops and Jamoytius are now considered more basal than the Anaspida proper. There is also some uncertainty whether these early types belong on the lamprey stem (diagram here) or are a seperate clade of early vertebrate evoluution (Sansom et al 2010a). The dendrogram given in Janvier 2007 p.30 differs slightly from the above, by placing hagfish, lampreys, Euphaneropids, possibly conodonts, and higher vertebrates in an unresolved polytomy. The status of the thelodonts is ambigious, they are variously placed above (Janvier 2007, Janvier 2008 fig 1) or below (Sansom et al 2010a) the Anaspida and Pteraspidomorphs. These latter two groups meanwhile have also played musical chairs, the Anaspida, previously more derived than the Pteraspidomorphi (Benton 2005 p.41) are now shown below them. However, whilst molecular phylogeny mostly argues for cyclostome monophyly (lampreys and hagfish sharing a distinct common ancestor separate from all other extant vertebrates - Delabre et al 2002 Kuraku et al 2009a, Kuraku et al 2009b, Heimberg et al 2010; with a few exceptions in older papers e.g. Gürsoy 2000) there is no morphological and paleontological evidence for such. Interestingly, Janvier, previously opposed this interpretation (Janvier 2008) now has come around in support of it (Janvier 2010). We will have more to say on this subject later (pending time constraints). For now we would tend to interprate unusual molecular phylogenies of this sort as methodological artifacts rather than empirical evidence of actual deep time phylogeny (similar instances include turtles as crown group archosaurs, and the monophyly of monotremes and marsupials against eutheria). This is yet another example of the need for a total evidence approach, rather then relying - as specialists inevitably tend to - on a single methodology to the exclusion of all others.
For now we have decided, rather unimaginitively, to go with the consensus approach of gnathostomes emerging from Cephalaspidomorphi, while retaining the thelodonts as a paraphyletic and possibily polyphyletic grouping of unarmoured agnaths on the way to the vertebrate condition (and sometimes evolving parallel to it)
Well-developed eyes; extrinsic eye muscles attached to the eyeball and orbital wall; 2-3 semi-circular canals; muscular, perforated pharynx; endoskeleton with at least cranium, visceral arches, limb girdles, and 2 pairs of appendages; repeating endoskeletal elements flanking the spinal cord, primitively two pairs in each metamere (interdorsals and basidorsals); radial muscles in fins; integument with two layers, epidermis and dermis; lateral line canals with true neuromasts; atrium and ventricle of heart closely-set; nervous regulation of heart by vagus nerve; blood with (a) red blood cells erythrocytes) containing hemoglobin, and leukocytes; mesodermal muscle fibers in gut; hox gene duplications; neural crest cells (arguably the basis for most other important vertebrate characters); sensory placodes (neuroectoderm discs giving rise to nose, lens of eye, ears and lateral line system).
checked ATW031116, revised MAK111027
