 Thoosuchinae: = Thoosuchidae) Thoosuchus, Prothoosuchus.
Thoosuchinae: = Thoosuchidae) Thoosuchus, Prothoosuchus.| Temnospondyli | ||
| The Vertebrates | Trematosauroidea |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Tetrapoda
├─┬─Lepospondyli
│ └─Reptiliomorpha
│
└─Temnospondyli
├─Edopoidea
└─┬─Dvinosauria
└─┬─Euskelia
│ ╞═Dissorophoidea
│ │ └─Lissamphibia
│ └─Eryopoidea
└─Stereospondyli
├─Rhinesuchidae
└─┬─Lydekkerinidae
├─┬─Plagiosauroidea
│ └─┬─Rhytidosteidae
│ └─Brachyopoidea
└─┬─Capitosauria
└─Trematosauria
├─Trematosauroidea
│ ├─Thoosuchinae
│ │ ├─Prothoosuchus
│ │ └─Thoosuchus
│ └─Trematosauridae
└─Metoposauroidea
|
Contents
Index |
According to Yates & Warren (2000), whom we have no reason to doubt, the middle part of the temnospondyl story goes something like this. Temnospondyls thrived throughout the Permian, during which they had a world-wide distribution. Whatever disaster overtook life on Earth at the end of the Permian killed off just about all of the temnospondyls in a fairly short space of time. Somewhere in Gondwana, a relict population held out. In the earliest Triassic (Induan), these forms radiated from Gondwana to repopulate the Earth. The two major branches of this renaissance were the Capitosauria e.g., lydekkerinids, capitosaurs, and mastodonsaurs) and the Trematosauria, including the Trematosauroidea and all other post-Permian temnospondyls.
Not surprisingly, some of the most basal temnospondyls we see are the trematosauroids. Its not surprising that we see them, because they are big and easy to find. Its not surprising that we see them early because they are the only stem tetrapods to have developed truly marine forms. We would thus anticipate that they would radiate fast and far. In fact, the odd thing is that we don't see them even earlier than the Oleneckian, some 2 My after the dawn of the Triassic.
At least this is what Damiani (2001) states. Damiani's paper is, in some ways, a paper of classic quality. In other respects, it is rather frustrating. In fairness, Damiani did not set out to study the Trematosauria. Quite the contrary. He aims at a comprehensive revision of the Capitosauria, or what he believes would be better termed the "Mastodonsauroidea." However, in order to delineate his group, he necessarily takes in some trematosaurian genera, including Thoosuchus, Angusaurus, and Trematosaurus. So far as the Capitosauria are concerned, he has certainly produced a definitive reorganization of the clade. However, some of the details relating to the taxa of interest to us on this page are unclear. See, for example, the comments set out as notes under the entries for Thoosuchusand Yarengia.
Another bone we would pick with Damiani relates to the bones he picked. That is, how can we possibly assess the phylogenetic position of the Capitosauria when the only other Triassic forms in the cladogram are trematosauroids? The problem can be summarized in the following, highly abbreviated, cladograms:
Yates & Warren (2000) |
Damiani (2001) |
|
Stereospondyli
├─Capitosauria
│ ├─Lydekkerinidae
│ └─┬─Mastodonsaurus
│ └─┬─Benthosuchus
│ └─Capitosauridae
└─Trematosauria
├─Trematosauroidea
└─┬─Metoposauroidea
└─┬─Plagiosauroidea
└─etc.
|
Stereospondyli
├─Lydekkerinidae
└─┬─Mastodonsauroidea
│ ├─Benthosuchus
│ └─Mastodonsaurus deeply nested in a mess of capitosaurids
└─Trematosauroidea
|
The differing position of the Lydekkerinids is obviously of some interest. To Yates & Warren, they are the sister of all Capitosauria. To Damiani, they end up being the sister group of all other Triassic temnospondyls. Benthosuchus is also of considerable importance. It has historically been regarded as a trematosaur of some kind and differs from them in only a few characters. Damiani asserts that it is the sister of all other capitosaurs and Mastodonsaurus. But how can we have any confidence in Damiani's findings about either group if we do not know the branch point leading to all other Triassic temnospondyls? And, if we are unsure about the positions of either lydekkerinids or Benthosuchus, how much confidence can we have in the rest of the arrangement?
At its most abstract -- but also most fundamental -- level, the problem is that you need a clade to make a cladogram. Unless Yates & Warren (and many others) are very far off base, the study taxa do not constitute a clade. Somewhere between the granddaddy stereospondyl and Trematosaurus there is a branch leading to hundreds of other species of stereospondyl, including metoposaurs, plagiosaurs, batrachosaurs and many, many others.
Absent a clade, it is a bit of a strain to talk about consistency, and that may be why Damiani does not use any tests of consistency. A consistency index measures how many times a character flips from one state to another in the cladogram. If a large chunk of the clade is missing, as here, there is no way to measure consistency. This is not simply a choice of statistical measures. The whole point of a parsimony-based cladogram is to be parsimonious -- that is, to minimize the total number of character state changes. That's not possible unless you have enough taxa to fairly sample the changes which occurred in the clade. If one arbitrarily hacks off branches, there is no valid measure of parsimony. We don't get a phylogenetic tree, but a paraphyletic topiary shaped by the choice of test taxa.
In the last analysis, we can confidently buy into Damiani's cladogram of the Capitosauridae, excluding Benthosuchus. This clearly leaves a monophyletic group, although it may unreasonably exclude lydekkerinids. This means that Damiani is likely correct in placing Mastodonsaurus deeply among the capitosaurids. Hence he may be correct in changing the name to Mastodonsauroidea. However, lacking valid information on the position of the two key sister taxa (lydekkerinids and Benthosuchus), it is more difficult to accept the higher-level placement of this clade. Since we can't yet be confident in the placement of Benthosuchus, it is too early to accept Mastodonsauroidea (= Benthosuchus + Mastodonsaurus) as a useful taxon. ATW020722.
Trematosauria: Trematosaurus > Parotosuchus.
Stereospondyli::: Capitosauria + *: (Trematosauroidea + Luzocephalidae) + (Metoposauroidea + (Plagiosauroidea + Rhytidosteidae + Brachyopoidea))).
Interorbital space > 50% of skull width at mid-orbit; foramen on stapes absent; no sharp rimmed depression on posteroventral surface of parasphenoid (reversal); denticle patches absent from coronoids.
References: Yates & Warren (2000). ATW000213.
Trematosauroidea: Thoosuchus + Trematosaurus [YW00] Vybrosaurus?
Early Triassic (Olneckian) [D01]
Phylogeny: Trematosauria:: Luzocephalidae + *: Thoosuchinae + (Yarengia + Trematosauridae).
Introduction: Large, gharial-like forms with elongated rostrums, probably specialized for catching fish. In keeping with Yates & Warren (2000) the short-lived northern Pangean Thoosuchinae (formerly considered basal capitosauroids) are transferred from the Capitosauroidea to the Trematosauroidea. They could perhaps be considered primitive members of this lineage.
Specialized long-nosed marine fish-eaters. Skull generally wedge-shaped, tall and narrow [S+00]; long snout formed by extended frontals, narials and premaxillae; nares long & narrow [D01]; orbits well-separated [S+00]; frontal excluded from orbit [S+00]; lateral projection of postorbital reduced or absent [S+00]; postorbital, prepineal growth zone present [D01] [1]; tabular horns and otic notch reduced or absent [S+00]; squamosal minimally embayed; sensory canal present on occiput [D01]; pterygoid oblique ridge reduced [S+00]; glenoid fossa below level of dorsal surface of dentary [D01]; $ parasphenoid plate extends posteriorly to cover exoccipital condyles in ventral ?probably means palatal) view [YW00]; $ parasphenoid cultriform process laterally compressed so that sides meet to form midline ventral keel [YW00][D01]; cultriform process underplated by a posterior extension of the vomers [D01]; pterygoid - parasphenoid suture extends back to underlie anterior part of middle ear cavity [S+00]; labial wall of adductor fossa arched dorsally (in lateral view) [D01]; ectopterygoid enters margin of interpterygoid vacuity [D01]; $ paired anterior palatal fossae [YW00] [S+00]; posterior Meckelian foramen long, exceeding 50% length of adductor fossa [D01]; pterygoid denticle fields well-developed [S+00]; ectopterygoid tusks absent in adults [D01] (contra, [S+00]); retain primitive paired pleurocentra in vertebrae; well-developed lateral line system [D01] [S+00]; at least some forms marine(!).
Notes: [1] Temnospondyls usually show a pattern of ornamentation on dermal skull bones consisting of a pattern of ridges and pits. In some cases, the pits seem to be stretched out into grooves. These are thought to represent regions of localized growth. A marked zone of this sort is characteristic of trematosauroids. Damiani (2001) lists it as a synapomorphy of the clade. However, Damiani did not include any trematosaurians in his study other than trematosauroids (i.e. no metoposaurs, plagiosaurs, rhytidosteids, etc.). As a result, the status of his trematosauroid synapomorphies is unclear. They may, instead, represent characteristics of all Trematosauria. Against the background of Yates & Warren (2000) it seems likely that Damiani is correct -- if nothing else, both Yates and Damiani did much of their critical work in Prof. Warren's lab at the same time. However, out of an abundance of caution, I have not flagged these characters as synapomorphies of Trematosauroidea on the basis of Damiani's study alone.
Links: Abstracts for PaleoBios 14(1-4) (include Lonchorhynchinae?); Trematosauroidea; Steyer (2002).
References: Carroll 1988); Damiani (2001) [D01]; Pough et al. (1999); Shishkin et al. (2000) [S+00]; Yates & Warren (2000) [YW00]. ATW030126.
Thoosuchinae: = Thoosuchidae) Thoosuchus, Prothoosuchus.
Early Triassic (Early Olneckian) [D01] of Russia.
Trematosauroidea: (Yarengia + Trematosauridae) + *: Prothoosuchus + Thoosuchus.
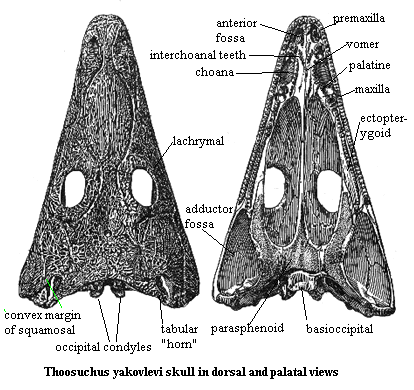
Characters: lacrimals present; orbits posterior to midpoint of skull; skull table not elongated; sensory sulci on skull roof; convex squamosal margin which bulges and narrows otic notch in ventral view [YW00]; deep otic notch; tabular well developed; tabular horns extend to posterolateral margin of skull; quadratojugal does not contribute to mandibular condyle [D01]; doubled occipital condyle, without contribution from basioccipital; anterior palatal fossae separated by median ridge; vomer, palatines, and maxilla all contribute to margin of choana; parasphenoid extends posteriorly to partially cover base of occipital condyles in ventral view (note that figure does not show this condition, & instead suggests that the basioccipital partially covers the condyles); some muscular crests on posterior of parasphenoid; premaxillary teeth unspecialized; vomer without denticles; ectopterygoid without tusks, with >3 rows of teeth; posterior coronoids with teeth (?); dermal bones ornamented with ridges enclosing pits.
Links: Thoosuchus; bone_clones_thoosuchus; Comparative Vertebrate Anatomy - Lecture Notes 1; TIKHVINSKOYE
Image: modified from More Thoosuchus yakovlevi, citing Ryabinin (1925).
Damiani (2001) [D01]; Yates & Warren (2000) [YW00].
ATW020719.
Prothoosuchus:Getmanov 1989. P. blomi Getmanov 1989, P. samariensis Getmanov 1989.
Range: Early Triassic of Russia.
Phylogeny: Thoosuchinae: Thoosuchus + *.
Characters: pineal foramen close to posterior margin of orbits [S+00]; parasphenoid cultriform process relatively broad [S+00].
References: Shishkin et al. 2000) [S+00]. ATW020720.
 Thoosuchus: Efremov 1940. T. yakovlevi Ryabinin 1927, T. tardus Getmanov 1989, T. tuberculatus Getmanov 1989.
Thoosuchus: Efremov 1940. T. yakovlevi Ryabinin 1927, T. tardus Getmanov 1989, T. tuberculatus Getmanov 1989.
Range: Early Triassic of Russia.
Phylogeny: Thoosuchinae: Prothoosuchus + *.
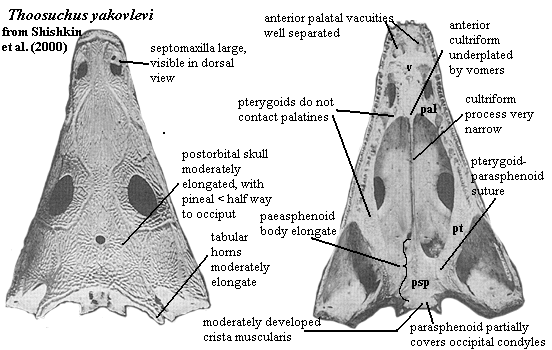
septomaxilla large and exposed on dorsal surface [S+00]; orbits broadly separated [S+00]; skull roof moderately elongated behind orbits [S+00]; pineal foramen in anterior 1/3 of orbital - occipital distance [S+00]; lateral projection of postorbital weak or absent [S+00]; posterior squamosal margin convex, narrowing otic notch [YW00] [1]; tabular horns moderately elongated [S+00]; parasphenoid plate elongate, with moderate cristae muscularis [S+00]; cultriform process very narrow [S+00]; pterygoid does not contact palatine [S+00]; anterior palatal vacuities broadly separated [S+00]; lower jaw with pronounced retroarticular process [S+00]; well-developed posterior Meckelian foramen [S+00]; surangular medial process weak [S+00]; tooth rows between choanae meet at acute angle [S+00].
Notes: [1] in Yates & Warren's (2000) study, this is the only character state which separates Thoosuchus from the Trematosauridae. Damiani 2001) states that they differ because Thoosuchus has a deep otic notch. Yates & Warren score this character as unknown for trematosaurids. Damiani also states that they differ because the occipital condyles in Thoosuchus are anterior to the quadrate condyles. In the image, this simply does not appear to be the case. However, compare the illustration of the same specimen from Ryabinin's original paper under Thoosuchinae.
Links: TIKHVINSKOYE; Thoosuchus; Thoosuchus Skull - Thoosuchus spp; TRIÁSSICO vertebrados (Spanish).
References: Damiani (2001); Shishkin et al. 2000) [S+00]; Yates & Warren (2000) [YW00]. ATW020720.
checked ATW050530
{kind=link}