Archosauria: Archosauria: Phytosauridae
Parasuchus & Paleorhinus
Taxa on This Page
- Paleorhinus X
- Parasuchus X
- Phytosauridae X
 |
|
A giant Pseudopalatus-grade phytosaur, "Rutiodon" lithodendrorum, length to 12 meters, startles a group of small fabrosaurs of the genus Revueltosaurus, on one of the wet floodplains of west equatorial Pangea (now, Petrified Forest National Park in Arizona), early Norian age. Illustration © Doug Henderson. |
The Phytosaurs
Introduction
The phytosaurs (this unfortunate name means "plant lizards", because it was mistakenly believed that petrified mud fillings in the jaw of the first specimen found were herbivore teeth) were crocodile-like semi-aquatic thecodonts that suddenly appeared and became very abundant during the latter part of the Triassic period.
They are more appropriately known by the later (but less widely used) name Parasuchia ("alongside the crocodiles"), as they resembled crocodiles to a remarkable degree in size, appearance, and life-style. This is an amazing illustration of convergent or parallel evolution. Phytosaurs were in a sense "crocodile uncles", since phytosaurs and proto-crocodiles shared a common ancestor among the early Crurotarsi. But true crocodiles evolved only long after phytosaurs became extinct at the end of the Triassic.
In some ways, the differences between phytosaurs and crocodiles are minor. The most obvious difference between phytosaurs and crocodiles is in the position of the nostrils. Phytosaurs had nostrils placed near or above the level of the eyes. Crocodiles have nostrils placed far forward at the tip of the snout. Another difference is in the palate (roof of the mouth). True crocodiles have a secondary palate that enables them to breathe when partially submerged, even though the mouth is full of water. Phytosaurs lacked this adaptation, and used the nostrils on top of the head in order to swallow air while underwater. Phytosaur limbs were also somewhat more primitive in structure than those of crocodiles. Fossil footprints indicate however that phytosaurs could move in a semi-erect stance on land and did not drag their tails as modern crocodiles do.
 |
|
Phytosaur dorsal scutes. |
Phytosaurs were if anything even better armoured than crocs, with the throat and back of the animal being are protected by heavy armoured scutes, and the belly reinforced with a dense arrangement of abdominal ribs. The armoured scutes are often found as fossils
Phytosaur Morphotypes and Life-style
In fact, phytosaurs are characterized by three distinctive cranial morphotypes, and this seems to be independent of phylogenetic relations. Modern crocodilians exhibit a similar morphological diversity. These skull patterns are linked to characteristics of the dentition; specifically the differentiation or similarity of the teeth along the jaws.
Dolichorostral ("long snouted") types have a long, narrow snout; and teeth that are homodont, or similar in form. Good examples include Paleorhinus, Rutiodon carolinensis, and Mystriosuchus, the latter representing the highest development. They were most likely piscivorous, like the Gavialidae. The long, narrow snout is suited for capturing fast prey, while the weak teeth could hold and position the fish for swallowing, but could not produced the shear needed to cut up chunks of flesh.
Altirostral ("high snouted") animals have an intermediate condition and primarily heterodont (different lengths or size) dentition. Typical forms might be Angistorhinus and Pseudopalatus. They were probably generalist feeders like the Alligatoridae.
Brachyrostral ("short snouted") forms have a massive, relatively broad snout, skull and jaws; and tends to extreme heterodont dentition. These animals most likely fed on large tetrapods. The mediolaterally flattened posterior teeth would slice the carcass into chunks suitable for swallowing. In addition, they probably used their jaws in intraspecific combat. Nicrosaurus and Smilosuchus are good examples of this morphotype; the latter representing the most extreme adaptation along these lines.
Phytosaur Biozonation.
Chatterjee, Lucas and others have shown that phytosaur genera can be used to identify the age of a particular formation. Phytosaurs are particularly useful for biozonation because these large river and swamp-dwelling animals were frequently fossilized. Thus Paleorhinus and the Angistorhinines dominated the middle (or early part of the late) Carnian. Rutiodon-like forms such as Leptosuchus and Smilosuchus (some of these names perhaps do not deserve distinct generic status - this is a classic instance of "splitters vs lumpers") become a key index fossil for the Adamanian interval (the later part of the late Carnian).
In a number of papers (e.g. Lucas 1998), Dr Spencer Lucas and co-workers have proposed a series of biozones characterized by distinctive genera for the Triassic. In each of the four biozones that make up the late Triassic, different phytosaur types are found. Possibly these intervals were separated by periods of extensive climate change, which greatly stressed these large semi-aquatic reptiles, leading to rapid speciation.
The following table shows each of the four Late Triassic biozones and representative phytosaur genera and subgenera for each:
| Geological Age |
Land Vertebrate Age |
Representative phytosaur genera and subgenera
|
Rhaetian
(also late Norian?)
|
Apachean |
Redondasaurus |

|
Middle Norian
Early Norian |
Revueltian |
Pseudopalatus, Nicrosaurus, Mystriosuchus.
|

|
| latest Carnian (= late Tuvalian)
|
Adamanian |
Rutiodon, Leptosuchus, and Smilosuchus |
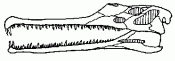 |
| earlier late Carnian (= early Tuvalian)
|
Otischalkian |
Paleorhinus, Angistorhinus |
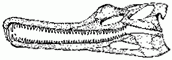
|
Classification
The classification of phytosaurs has been somewhat debated. Gregory 1969 (cited in Westphal, 1976 p.117) has three separate lineages, the slender-snouted Paleorhinus-Mystriosuchus, the more heavily built Angistorhinus-Rutiodon/Belodon, and the most extreme (crested snouted) Nicrosaurus.
Chatterjee has only two families, the Parasuchinae, which includes not only primitive types such as Parasuchus and Paleorhinus, but also the typical forms like Rutiodon and Nicrosaurus and the Mystriosuchinae for Francosuchus and Mystriosuchus
The traditional view since then has the phytosaurs divided into two subfamilies (recently considered distinct families): the slender-snouted gavial-like Mystriosuchinae (including Paleorhinus) and the heavy crocodile or alligator-snouted Angistorhininae (= Rutiodontidae). The former would seem to have fed largely on fish and perhaps other small animals, the latter on larger prey as well as fish. Both appear suddenly in the Middle Carnian and continue through to the very end of the Triassic period.
Advocates of this approach to phytosaur classification (e.g. Long & Murry 1995) tend to divide the group into two distinct (but related) families, as follows:
Parasuchia Huxley 1875
Mystriosuchidae Huene, 1915
Palaeorhininae Long & Murry, 1995
Mystriosuchinae Nopcsa, 1928
Angistorhinidae Huene, 1936 (= Rutiodontidae Long & Murry, 1995)
Angistorhininae Camp, 1930
Rutiodontinae Long & Murry, 1995
Pseudopalatinae Long & Murry, 1995
Of the above taxa, the Palaeorhininae and the Angistorhininae are the two oldest groups; these are replaced in the Later Carnian by the Rutiodontinae. The Norian then saw these in turn replaced by the Pseudopalatinae and the Mystriosuchinae. The Pseudopalatinae survived till the end of the Rhaetian.
In an alternative classification [Doyle & Sues, 1995, cited in Mikko's Phylogeny - Phytosauria (Parasuchia)], the palaeorhinines (as represented by the genus Paleorhinus) are considered the primitive sister group of all other phytosaurs. The similarity between Paleorhinus and Mystriosuchus are thus the result of parallel evolution, not direct ancestry and descent. This is quite likely, considering that no intermediate forms between Paleorhinus and Mystriosuchus are known in the latest Carnian or the early Norian; such a long gap in animals so well represented in the fossil record is unusual.
A third approach, indicated by biozonation in association with plesiomorphic or highly derived traits, has the early, unspecialized, forms like Parasuchus giving rise to the Angistorhinidae, with the paraphyletic/grade-level Angistorhininae in turn being replaced by their descendents the rutiodontine-grade phytosaurs in the Later Carnian, and these in turn giving rise to the most advanced clade, the Pseudopalatinae, which unlike the earlier taxa is a proper monophyletic grouping, in the Early Norian. This succession of early primitive types with later more advanced forms, and those with their still more advanced descendents, is not unlike the evolution of equids, titanotheres, and other fast-evolving taxa. Of course real-world evolution is more an unruly bush than a tidy ladder, but the generic pattern could still hold to some extent.
In view of no agreed-upon model of phytosaur evolution, the following phylogeny is proposed, based on a synthesis of a number of approaches, along with the author's rampant speculation, and thus should not be considered definitive. (MAK 030727)
Descriptions
 Phytosauridae:(= Parasuchia): Angistorhinus, Mystriosuchus, Nicrosaurus, Paleorhinus, Rutiodon
Phytosauridae:(= Parasuchia): Angistorhinus, Mystriosuchus, Nicrosaurus, Paleorhinus, Rutiodon
Late Triassic of Europe, India, North America, North Africa (Morocco), Madagascar(?), China(?).
Characters: 3-12 m; generally similar to crocs in form & niche; skull length >50% of length of presacral vertebral column [$π03]; skull dorsoventrally compressed [S91]; premaxilla extended [$S91]; piscivorous snout [$S91]; "septomaxilla" (neomorph?) with broad dorsal exposure, sutured to antimere at midline [$S91]; nasal with ventrolateral process extending anterior to nares [$S91]; narial openings far back on snout & elevated above orbits; prefrontal, descending process absent [π03]; nares & orbits dorsally placed [$S91]; quadratojugal generally triangular [$S91]; occiput twice as wide as tall [$S91]; no bony secondary palate; palatine forms entire lateral margin of choana & extends anteriorly to contact premaxilla [$S91]; palatine with longitudinal ridge (margin of soft secondary palate) [$S91]; postpalatine fenestra reduced to a slit [$S91]; foramina for cebreral branch of internal carotids on posterolateral corner of basisphenoid [$G02] [$π03]; lateral wall of braincase formed by large laterosphenoid (also aetosaurs & crocs); some with fang-like teeth in expanded symphysial area of both jaws; gastralia present; interclavicle enlarged [$S91]; coracoid crecentic [$S91]; forelimbs slightly shorter than the hind; grasping manus?; large, spreading feet & hands; manus V with <4 phalanges [$π03]; pubis & ischium plate-like, primitive; fifth metatarsal hooked & calcaneal tuber lateral primitive leverage); throat & back with scutes; paired paramedian osteoderms, with small lateral osteoderms intercalated between lateral tips of main plates [S92]; osteoderms sculptured [$π03]; semi-erect trackway evidence; lived near fresh water (no known marine species).
Notes: "[π03]" refers to our own analysis of published data.
Links:
Phytosaur; dinopics; A PRELIMINARY BIOMECHANICAL; Phytosauria Translation and Pronunciation Guide Introduction; Walking with Dinosaurs - Fact Files; Phytosaur - Rutiodon; Lecture 10 - Triassic: Newark, Chinle; Dino Land Travels Database Dinosaur State Park: Rutidon Model; Rutiodon; Phytosauria [Parasuchia]; Habay-la-Vielle 2 (French).
References: Chatterjee (1986); Gower (2002) [G02]; Long & Murry (1995), Sereno (1991) [S91], Sues (1992) [S92]. MAK030812, ATW031226.
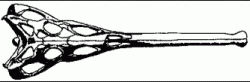 |
|
Skull of Parasuchus hislopi; length about 60 cm.
Source: Westphal, 1976, p.115. |
Parasuchus: Parasuchus hislopi
Range: Late Triassic (earlier Late Carnian) of India.
Phylogeny: Phytosauridae : Paleorhinus + Euphytosauridae + *.
Characters: Small, skull-length about 60 cm; moderately deep skull, orbit directed outwards and upwards; rostrum slender and without crest; nostrils forward along the snout, i.e. external nares well in front of the antorbital fenestrae, squamosals without any posterior process; supratemporal fenestrae at the level of the skull roof, large interpterygoid vacuity, (like Paleorhinus); posttemporal fenestrae moderately sized; teeth short and slender, weakly heterodont; overlapping scutes; mildly dolichorostral.
Comments: This animal is more usually included under Paleorhinus. As would be expected, there is a story behind the nomenclatural ambiguity. The original description was by Thomas Huxley (who made a name for himself as "Darwin's Bulldog") based on rather fragmentary materials, which another early paleontologist, Lydekker, further confused by including rhynchosaur material. Because not much was known about this animal, similar animals discovered elsewhere were given new generic names. In 1978 Chatterjee assigned two nearly complete specimens from the Maleri Formation to Parasuchus hislopi. Since the Indian Parasuchus and North American Paleorhinus differ in only the most minor details, the latter becomes a junior synonym of the former. Later researchers have preferred to identify the new Indian specimens as a distinct species of the well-known Paleorhinus, and have restricted the genus Parasuchus to Huxley's type material as a nomen dubium (not enough material to give a definitive definition as a type species), although Chatterjee (1974) took the position that the type species was diagnosable based on a portion of a jaw from Huxley's material. [Creisler 1996). Although it is not impossible they represent a separate species, it is much less likely that Chatterjee's specimens represent a distinct genus to Huxley's, especially since all palaeorhinines are currently included under a single genus.
Considered by Chatterjee a subgenus, although other sources simply sink it into Paleorhinus. P. hislopi possesses some characteristics such as the weakly heterodont teeth, that indicate it may be more plesiomorphic (closer to the common ancestor) than Paleorhinus proper. In all likelihood, Paleorhinus may turn out to be a paraphyletic assemblage of early phytosaurs.
References: Chatterjee (1978); Creisler 1996. MAK030812.
 |
|
Skull of Paleorhinus bransomi; length about 80 cm.
Source: A. H. Müller, Lehrbuch der Paläozoologie. |
Paleorhinus: Paleorhinus bransomi, Paleorhinus fraasi (formerly Francosuchus), Paleorhinus ehlersi (formerly Promystriosuchus)
Range: Late Triassic (Middle and earlier Late Carnian) of Europe, North America, North Africa (Morocco).
Characters: low skull, orbit directed more upwards than outwards; Small to medium-sized, skull-length 60-90 cm; rostrum slender to very slender; as in Parasuchus the external nares well in front of the antorbital fenestrae, squamosals without any posterior process, and supratemporal fenestrae at the level of the skull roof; posttemporal fenestrae small; teeth homodont; non-overlapping scutes; large interpterygoid vacuity; teeth short and slender, homodont; dolichorostral morphotype.
Comments: Considered by Chatterjee a subgenus of Parasuchus. He also considers Francosuchus a distinct genus, although in his short genus-level review of the family it is almost identical to Paleorhinus (but not to Parasuchus, hence I have retained a distinction between the two latter). The subfamily Palaeorhininae is probably a plesiomorphic grade, united by shared primitive characteristics, with more advanced phytosaurs separating at an early stage,
Links: Late Triassic graveyard of large Triassic tetrapods in the Opole Silesia (in Polish);
References: Chatterjee (1978). MAK030812.
checked ATW050702, last modified MAK050119
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners
