
Eryops at the Münchehagen Dino-Park, photo by GermanOle (Wikipedia, GNU Free Documentation License)
| Temnospondyli | ||
| The Vertebrates | Temnospondyli: Eryopoidea |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Tetrapoda
├─┬─Lepospondyli
│ └─Reptiliomorpha
│
└─Temnospondylii
├─Edopoidea
└─┬─Dvinosauria
└─┬─Euskelia
│ ╞═Dissorophoidea
│ │ └─Lissamphibia
│ └─Eryopoidea
└─Stereospondyli
├─Rhinesuchidae
└─┬─Lydekkerinidae
├─┬─Plagiosauroidea
│ └─┬─Rhytidosteidae
│ └─Brachyopoidea
└─┬─Capitosauria
└─Trematosauria
├─Trematosauroidea
└─Metoposauroidea
|
Contents
Index |
|
Eryops at the Münchehagen Dino-Park, photo by GermanOle (Wikipedia, GNU Free Documentation License) |
 Eryops skeleton. Photo by Joshua Sherurcij © 2007 |
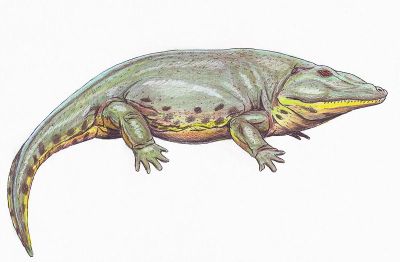
The classical temnospondyl is Eryops (right): a big, flat slow-looking tetrapod with an enormous mouth. The classical definition of a temnospondyl involved vertebral characters. That is, the principal vertebral component was the intercentrum which (to varying degrees) tended to grow up and around the notochord and provided the basic structural support for the axial skeleton. By contrast, in reptilomorphs (i.e. amniotes and their immediate ancestors), the pleurocentrum grows down over the notochord, fuses with the neural arches, and eventually drives the intercentrum into anatomical oblivion. Actually, the intercentrum of Eryops is not all that well developed, even compared to Ichthyostega. Nevertheless, Eryops is the archetype around which the idea of a temnospondyl revolves.
Like most Carboniferous and Permian tetrapods, Eryops is a cipher. Even on a page devoted to speculation, it is difficult to find a way to fit these parts together. For example, it is all well and good to say that it used an inertial snap of the jaws to capture prey; but it is difficult to snap anything that big underwater, and equally difficult to imagine Eryops as a terrestrial hunter. Nevertheless, the dentition is rather unambiguous. It ate relatively large things that were not happy about the idea. The mouth appears to be well-designed for swallowing animals more or less whole and for keeping them there, in spite of strenuous objections, with such devices as internal fangs.
Perhaps the back forms a sort of an arch, peaking in front of the tall pelvis. This would bring the intercentra into relatively close contact, supporting the arch from below. The neural arches are not smooth curves, as in Ichthyostega. Instead, they are strongly sculpted, working tools for complex attachments of tendon and muscle. One could imagine a style of movement or predation in which substantial energy is stored in arching the back by movement of the hind legs, and then releasing it - perhaps quite suddenly - by stepping forward with the arms and contracting muscles anchored on the dorsal spine. This may be consistent with the odd pattern of early "tetrapod" tracks found in Nova Scotia as well as the structure of the ribs, which appear capable of sliding past each other. ATW000213.
Range: from the Late Carboniferous.
Phylogeny: Temnospondyli::: Limnarchia + *. Eryopoidea + Dissorophoidea.
Some large, wide forms up to 2 m. Description is largely of Eryops. Large, wide skull; skull shape generally U-shaped; lower jaw triangular in lateral view; long, sharp labyrinthodont teeth; some accessory fangs on ectopterygoids, palatine, vomers, etc. (probably used inertial snap of jaws); nares almost terminal; depth of skull increases posteriorly, with fairly pronounced upward curvature ant to orbits; orbits face antero-laterally variable); dermal skull ornamented with pits; $ intertemporal absent; $ parasphenoid firmly attached (sutured) to pterygoid, with no moving articulation between braincase and pterygoid; no separate cervical vertebral series (no neck!); dorsal vertebrae with moderately tall, thick neural arch; both pleurocentrum and intercentrum rather small; ribs broad pronounced in Eryops); ribs shorten post, and may be absent at level of sacrum; 1 sacral, not (or not completely?) fused to ilia; caudal ribs variable in number and morphology, but tail is not longer than dorsal series; pectoral girdle not attached to skull; massive scapula; coracoid expanding to plate ventro-medial to glenoid; ilia oddly tall; humerus and femur short and massive; humerus & tibia oriented horizontally; radius & ulna short and well-separated, as are tibia & fibula; 3 carpals; 5 (4?) digits on manus, 5 on pes; Branchiosaurs may be embryonic and/or neotenous euskelians with external gills.
Links: The Field Museum of Natural History Eryops; subcl98 labyrinthodont teeth, link to sketch); eryops dorsal skull); permian image page (life, beautiful!); Euskelia Mikko's phylogeny). ATW030129
Early to Late Permian of North America & Europe.
Euskelia: Dissorophoidea + *.
Characters: choana relatively rounded; iliac blade vertical.
Note: The Eryopoidea were for a long time a sort of waste-basket taxon for mostly late Carboniferous and Early Permian temnospondyls that could not be slotted anywhere else. Yates & Warren (2000) reduce this to two families, the Eryopidae and Zatracheidae, although other families may also belong here.
Image: Eryops image courtesy of Crash Jones. ATW020213.
Links: Temnospondyli; Eryopoidea; JOINT ADVENTURE Ulla Lohmann; Best on the Web, also in German. The figures are included in the German text); Phylogeny and Apomorphies of Temnospondyls ToL). ATW030616.
Timothy Pilgrim - from the Wikipedia history log of Eryops
Note by editor - a useful essay (although using pre-cladistic terminology) that appeared on the Wikipedia Eryops page before being successively whittled away. I thought it worth including in its entirety (The images were added to later revisions of the page). - MAK090723
Introduction
Two hundred eighty million years ago marked the beginning of the Permian period in the history of the Earth. The continents of Laurasia and Gondwana were coming together, forming the supercontinent Pangea. The waters were populated with fish and invertebrates, and the land-bound margins were swampy areas, covered by dense vegetation. These swamps are of significant importance to natural history, for they were the residence of the first tetrapods, who had evolved beyond lobe-finned fish into air-breathing, quadrapedal animals, designated the amphibians.
 Eryops megacephalus - artwork by Dmitry Bogdanov, wikipedia |
Aside from the fact that they gave rise to the reptiles, the amphibians are an intriguing group in their own. One such animal was Eryops, a six-foot long, semi-aquatic belly-dragger found in the Wichita and Clear Fork deposits of Texas and New Mexico. Additional fossils have also been found in the eastern United States. Although it gave no evolutionary progeny of its own, Eryops is the most well known Permian amphibian, and a remarkable example of natural engineering.
The transition from an aquatic lobe-finned fish to a quadrapedal air-breathing amphibian was a momentous occasion in the natural history of the vertebrates. For an animal to live in a gravity negligible, aqueous environment, then invade one which is entirely different required major changes to the overall body, in form and in function. Eryops is an example, one of many, of such an adaptation. It retained, and refined, most of the traits found in its fish ancestors. Sturdy limbs supported and transported its body while out of water. A thicker, stronger backbone prevented its body from sagging under it own weight. And by utilizing available fish jaw bones, a rudimentary ear was developed allowing Eryops to hear new airborne sounds.
Ancestry
Amphibians arose from a piscian ancestor since fish were the only existing vertebrates before the invasion of the tetrapods. Such a fish must have possessed similar traits to that of the early amphibians and then passed them on, including internal nostrils (to separate the breathing and feeding passages) and a large fleshy fin which could give rise to the tetrapod limb. It was the rhipidistian crossopterygians which fulfilled every requirement for ancestry. Their palatal and jaw structures were identical to amphibians and the dentition was identical, with labyrinthine teeth fitting in a pit-and-tooth arrangement on the palate. The crossopterygian paired fins were smaller than tetrapod limbs, but the skeletal structure was very similar in that the crossopterygian had a single proximal bone (analogous to the humerus or femur), two bones in the next segment (forearm or lower leg), and an irregular subdivision of the fin, roughly comparable to the structure of the carpus / tarsus and phalanges.
The major difference between crossopterygians and amphibians was in relative development of front and back skull portions; the snout is much less developed than in most amphibians and the post orbital skull is exceptionally longer than an amphibian's.
A great many of the early amphibians lived during the Carboniferous, therefore, an their ancestor would have lived earlier, during the Devonian. Ichthyostegids were the earliest of amphibians with a skeleton directly comparable to that of rhipidistian ancestors. Early Labyrinthodonts (Late Devonian to Early Mississippian) still had some ichthyostegid features such as similar skull bone patterns, labyrinthine tooth structure, the fish skull-hinge, pieces of gill structure between the cheek & shoulder, and the vertebral column. They had, however, lost several other fish features such as the fin rays in the tail.
Tetrapod Origin
Early amphibians undoubtedly lived in near-land areas of water. If the water had dried up, any fish would have become stranded and soon die, but amphibians could move (albeit cumbersome) to the next water hole and continue to exist. Some may have lingered around the newly formed mudhole and fed on stranded fish; others may have remained on land and eaten smaller amphibians, eventually establishing a true terrestrial fauna.
In order to propagate in the terrestrial environment, certain challenges were to be overcome. The animal's body needed additional support because buoancy was no longer a factor. A new method of respiration was required in order to extract atmospheric oxygen instead of oxygen dissolved in water. A means of locomotion would need to be developed to traverse great distances between waterholes with little difficulty. Water retention was now important since it was no longer the living matrix, and it could be lost easily to the environment. Finally, new sensory input systems were required if the animal was to have any ability to function reasonably while on land.
Classification
 Front view of Eryops megacephalus |
Diagnostic features unique to the Labyrinthodontia are hard to find at first glance; the complex dentine infolding tooth structure was shared with crossopterygian fish. The labyrinthodonts are divided into the Temnospondyli and the Anthracosauria, the main difference between the two groups being their respective vertebral structures. The Anthracosauria had small pleurocentra which grew and fused, becoming the true centrum in vertebrates higher than themselves. In contrast, the Temnospondyli had a conservative vertebral column in which the pleurocentra remained small in primitive forms, vanishing entirely in the more advanced ones. The intercentra are large and form a complete ring.
A diagnostic feature of the Temnospondyli was that the tabular bone in the skull roof is relatively small and had no contact with the parietal, whereas contact between the two bones was present in all anthracosaurs.
Although the temnospondyls flourished in many forms in the Late Palaeozoic and Triassic, they were an entirely self-contained group and did not give rise to any later tetrapod groups. It was the sister group Anthracosauria which gave rise to the reptiles.
Within the Temnospondyli are the two suborders Rachitomi and Stereospondyli, also distinguished by their types of vertebrae. There were three distinct successive stages within the Rachitomi, the first occuring in the Carboniferous. The second happened mostly in the Pennsylvanian, continuing into Permian, of which Edops is characteristic. The third and final stage was in the Late Carboniferous and Early Permian, from which Eryops of the Texas Permian red beds is best known. Just as there were numerous side branches throughout the evolution of the temnospondyls, so too were there many of the rachitomes; Eryops represents but only one.
 Pelosaurus laticeps, a possible tadpole of Eryops (reference) |
Of special interest in regards to the Rachitomi, is Branchiosaurus. This tiny amphibian, relatively speaking, lived from the Late Pennsylvanian to the Early Permian and was very similar to the Rachitomi, differing only in its small size. However, it had a much less ossified skeleton, a short skull and other distinguishing features. Clear traces of gills are present in many fossilized samples, hence the name. Thought to have differed from rachitomous vertebrae, it was placed in a separate order named Phyllospondyli. Only later was it realized, by studying growth stages and seeing increasing ossification in larger specimens, that it was in fact the larval stage of a much larger rachitome like Eryops.
Backbone
There are two major forms of vertebrae found within the Amphibia. The first is the lepospondylous (husk) vertebra which is found in numerous small Palaeozoic forms as well as in modern amphibians. The second type is the labyrinthodont (arch) vertebra which occured in labyrinthodonts and, in a modified form, in all higher vertebrates. The latter type is a direct inheritance from the Crossopterygia, with the intercentrum and the pleurocentrum present.
Of the labyrinthodont vertebra, there are two subdivisions. Members of the Stereospondyli had intercentra which grew upwards, forming a ring about the notochord, and lacked any pleurocentra. The second, of which Eryops is an example, is the rachitomous-type vertebra which is the most primitive arch of all tetrapods. The intercentrum was a median ventral element encircling the persistant notochord, becoming the major element in construction of the centrum. The pleurocentra, which were small paired blocks flanking inbetween the intercentrum and neural arch, combine with the intercentum to make up the true centrum in higher vertebrates. The rachitomous vertebra is present in the oldest of amphibians, Ichthyostega, which were nearly identical to those of Crossopterygia, and the majority of Temnospondyli. The smaller pleurocentra were well ossified and occupied the space between the successive intercentra, making a vertebral column which was well built to support the weight of a terrestrial body. The development of anterior and posterior zygopophyses was an envolutionary necessity for tetrapods in order to strengthen the spine and control its flexibility. Eryops, like the majority of amphibians, had a single sacral rib with a broad head. This wielded a large capitulum and tuberculum, a short broad neck, and an expanded spatulate shaft. In addition, amphibians were the first vertebrates to have developed a neck which contained a more flexible cranio-vertebral joint than that of their ancestors.
Skull
 Side view of Eryops megacephalus |
The most notable characteristic that is different between a fish and amphibian skull are the relative frontal and rear portion lengths. The fish had a long rear portion while the front was short; the orbital vacuities were thus located towards the anterior end. In the amphibian, the front of the skull lengthened, positioning the orbits farther back on the skull. The lacrimal bone was not in contact with the frontal anymore, having been separated from it by the prefrontal bone. Also of importance is that the skull was now free to rotate from side to side, independent of the spine, on the newly forming neck. The average length of an Eryops skull was between 320 mm and 335 mm.
As with the flattening body, the skull of Eryops was dorsal-ventrally compressed, with a pitted surface of about 25 pits per square inch. The eyes which were originally on the sides, facing outward, were necessarily on the dorsal side of the skull looking up and forward. Although ossified in crossopterygians and ichthyostegids, the braincase of Eryops was not, in the orbital and otic regions. At the rear of the skull the once single occipital condoyle began to divide into two, but never fully separated.
A diagnostic character of temnospondyls is that the tabular bones (which formed the posterior corners of the skull-table) were separated from the respective left and right parietals by a sutural junction between the postparietals and supratemporals. Also at the rear of the skull, all bones dorsal to the cleithrum were lost.
The lower jaw of Eryops resembled its crossopterygian ancestors in that on the outer surface lay a long dentary which bore teeth. There were also bones below the dentary on the jaw: two splenials, the angulary and the surangular. On the inside were usually three coronoids which bore teeth and lay close to the dentary. On the upper jaw was a row of marginal labyrinthine teeth, located on the maxilla and premaxilla. In Eryops, like all early amphibians, the teeth were replaced in waves which traveled from the front of the jaw to the back in such a way that every other tooth was mature, and the ones in between were young.
Dentition
The Labyrinthodontia had a peculiar tooth structure from which their name was derived and, although not exculsive to the group, the labyrinthine dentition is a useful indicator as to proper classification. The important feature of the tooth is that the enamel and dentine were folded in such a way as to form a complicated corrugated pattern when viewed in cross section. This infolding resulted in strengthening of the tooth and increased wear resistance. Such teeth survived for 100 Ma, first among crossopterygian fish, then stem reptiles. Modern amphibians no longer have this type of dentition but rather pleurodont teeth, in fewer numbers.
Sensory Organs
There is a density difference between air and water that causes smells (certain chemical compounds detectable by chemoreceptors) to behave differently. An animal first venturing out onto land would have difficulty in locating such chemical signals if its sensory apparatus was designed for aquatic detection.
Fish have a lateral line system which detects pressure fluctuations in the water. Such pressure is non-detectable in air, but grooves for the lateral line sense organs were found on the skull of labyrinthodonts, suggesting a partially aquatic habitat. Modern amphibians, which are semi-aquatic, exhibit this feature whereas it has been retired by the higher vertebrates. The olfactory epithelium would also have to be modified in order to detect airborne odours.
In addition to the lateral line organ system, the eye had to change as well. This change came about because the refractive index of light differs between air and water, so the focal length of the lens was altered in order to properly function. The eye was now exposed to a relatively dry environment rather than being bathed by water, so eyelids developed and tear ducts evolved to produce a liquid, moistening the eyeball.
Hearing
The balancing function of the middle ear was retained from the fish ancestory, but delicate air vibrations could not set up pulsations through the skull in order for it to function a proper auditory organ. Typical of most labyrinthodonts, the spiracular gill pouch was retained as the otic notch, closed in by the tympanum, a thin, tight membrane.
The hyomandibula of fish migrated upwards from its jaw supporting position, and was reduced in size to form the stapes. Situated between the tympanum and braincase in an air-filled cavity, the stapes was now capable of transmitting vibrations from the exterior of the head to the interior . Thus the stapes became an important element in an impedance matching system, coupling airborne sound waves to the receptor system of the inner ear. This system had evolved independantly within several different amphibian lineages.
In order for the impedance matching ear to work, certain conditions had to be met. The stapes must have been perpendicular to the tympanum, small and light enough to reduce its inertia and suspended in an air-filled cavity. In modern species which are sensitive to over 1 kHz frequencies, the footplate of the stapes is 1/20th the area of the tympanum. However, in early amphibians the stapes was too large, making the footplate area oversized, preventing the hearing of high frequencies. So it appears that only high intensity, low frequency sounds could be detected, with the stapes more probably being used to support the braincase against the cheek.
Girdles
The pectoral girdle of Eryops was highly developed with a larger size for an increased muscle attachment to it and to the limb. Most notably, the shoulder girdle was disconnected from the skull resulting in improved terrestrial locomotion. The crossopterygian cleithrum was retained as the clavicle, and the interclavicle was well developed, lying on the underside of the chest. In primitive forms, the two clavicles and the interclavical could have grown ventrally in such a way as to form a broad chest plate, although such was not the case in Eryops. The upper portion of the girdle had a flat, scapular blade with the glenoid cavity situated below performing as the articulation surface for the humerus, while ventrally there was a large, flat coracoid plate turning in toward the midline.
The pelvic girdle also was much larger than the simple plate found in fishes, accomodating more muscles. It extended far dorsally and was joined to the backbone by one or more specialized sacral ribs. The hind legs were somewhat specialized in that they not only supported the weight, but also provided propulsion. The dorsal extension of the pelvis was the ilium, while the broad ventral plate was comprised of the pubis in front and the ischium in behind. The three bones met at a single point in the center of the pelvic triangle called the acetabulum, providing a surface of articulation for the femur.
The main strength of the ilio-sacral attachment of Eryops was by ligaments, a condition structurally, but not phylogenetically, intermediate between that of the most primitive embolomerous amphibians and early reptiles. The condition which is more usually found in higher vertebrates is that cartilage and fusion of the sacral ribs to the blade of the ilium are utilized in addition to ligamentous attachments.
Limbs
The humerus was the largest bone of the arm, its head articulating with the glenoid cavity of the pectoral girdle, distally with the radius and ulna. The radius resided on the inner side of the forearm and rested directly under the humerus, supporting much of the weight, while the ulna was located to the outside of the humerus. The ulna had a head, which muscles pulled on to extend the limb, called the olecranon that extended above the edge of the humerus.
The radius and the ulna articulated with the carpus which was a proximal row of three elements: the radiale underlying the radius, the ulnare underneath the ulna and an intermedium between the two. A large central element was beneath the last and may have articulated with the radius. There were also three smaller centralia lying to the radial side. Opposite the head of each toe lay a series of five distal carpals. Each digit had a first segment, the metacarpal, lying in the palm region.
The pelvic limb bones were essentially the same as in the pectoral limb, but with different names. The analogue to the humerus was the femur which was longer and slimmer. The two lower arm bones corresponded to the tibia and fibula of the hind leg, the former being the innermost and the latter the outermost bones. The tarsus is the hind version of the carpus and its bones correspond as well.
Feeding
Early amphibians like Eryops had a wide, gaping jaw with weak muscles with which to open and close it. Within the jaw were fang-like palatal teeth which, when coupled with the gape, suggests an intertial feeding habit. This is when the amphibian would grasp the prey and, lacking any chewing mechanism, toss the head up and backwards, throwing the prey farther back into the mouth. Such feeding is seen today in the crocodile and alligator.
The tongue of modern adult amphibians is quite fleshy and attached to the front of the lower jaw, so it is reasonable to speculate that it was fastened in a similar fashion in primitive forms, although it was probably not specialized like it is in a frog.
It is taken that Eryops was not very active, thus a predatory lifestyle was probably not the norm. It is more likely that it fed on fish either in the water or on those which became stranded at the margins of lakes and swamps. Also abundant at the time was a large supply of terrestrial invertebrates which may have provided a fairly adequate food supply.
Respiration
Modern amphibians breathe by inhaling air into their lungs, where oxygen is absorbed, as well as through the moist lining of the mouth and skin. So too did Eryops, but its ribs were too closely spaced to suggest that it simply expanded the rib cage. More likely, it depressed the hyoid apparatus to expand the oral cavity, and elevated the floor of the mouth while it and the nostrils were closed, forcing air back into the lungs. Air could then be forced back out by contraction of the elastic tissue in the lung walls. The other special respiratory methods were probably also made use of.
Locomotion
Eryops had typical amphibian posture exhibited by the upper arm and upper leg extending nearly straight out from its body while the forearm and the lower leg extended downward from the upper segment at a near right angle. The body weight was not centered over the tops of the limbs, but rather was transfered 90 degrees outwards and down through the lower limbs, which contacted the ground. Most of the animal's strength was used to just elevate the body off the ground in order to walk which was slow and difficult. With this sort of posture, only short, broad strides could be achieved, and have been so confirmed by fossilized footprints found in Carboniferous rocks.
Ligamentous attachments within the limbs were present in Eryops, being important because they were the precursor to bony and cartilagenous variations seen in modern terrestrial animals which use their limbs as a means of transport.
Of all body parts, the spine was the most affected with the move from water to land. It now had to resist the bending caused by body weight, and had to provide mobility where needed; previously, it was able to bend along its entire length. Likewise, the paired appendages had not been formerly related to the spine, but the slowly strengthening limbs now transmitted their support to the axis of the body.
Timothy Pilgrim, Gnu open source/Creative Commons license, 4 February 2004
checked ATW060327

{kind=link}
{kind=link}
{kind=link}