 Eoherpeton: E. watsoni Panchen 19??
Eoherpeton: E. watsoni Panchen 19??| Reptiliomorpha | ||
| The Vertebrates | Reptiliomorpha |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Tetrapoda
├─Temnospondyli
└─┬─Lepospondyli
└─Reptiliomorpha
├─Anthracosauroidea
│ └─┬─Eoherpeton
│ ├─Embolomeri
│ └─Chroniosuchia
└─┬─Gephyrostegidae
└─Batrachosauria
├─Seymouriamorpha
└─┬─Solenodonsaurus
└─Cotylosauria
├─Diadectomorpha
└─Amniota
|
Contents
Reptiliomorpha |
During the Carboniferous and Permian periods, prehistoric amphibians evolved along a number of parallel lines towards a reptilian condition. Some of these amphibians (e.g. Archeria, Eogyrinus) were elongate, eel-like aquatic forms with diminutive limbs, while others (e.g. Seymouria, Solenodonsaurus, Diadectes, Limnosceles) were so reptile-like that until quite recently they actually have been considered reptiles, that some of them they may still turn out to be reptiles, and it is likely that to an observer they would have appeared indistinguishable from small or large reptiles.
Although the first amniote reptile probably appeared in the late or even middle Mississippian period (Middle Carboniferous), as forms like Casineria would seem to indicate, a number of lines of reptiliomorph amphibians and proto-amniotes continued to flourish alongside their fully reptilian (crown amniote, to use the cladistic term) descendents and relatives for many millions of years.
By the middle Permian these various terrestrial forms had died out, but several aquatic groups continued to the end of the Permian, and in the case of the Chroniosuchids survived the end Permian mass extinction, only to die out at the end of the Middle or even the early Late Triassic. Meanwhile, the single most sucessful daughter-clade of the Reptiliomorphs, the Amniotes, continued to flourish and to inherit the Earth MAK050702 (for Wikipedia), MAK111108
The name Reptiliomorpha was coined by Professor Gunnar Säve-Söderbergh in 1934 to designate various types of late Paleozoic reptile-like labyrinthodont amphibians. (Säve-Söderbergh 1934) However Alfred Sherwood Romer used the name Anthracosauria instead. In 1970, the German paleontologist Panchen reverted to Säve-Söderberghs definition, (Panchen 1970) but Romer's terminology is still in use, e.g. Carroll 1988 and 2002, and Hildebrand & Goslow 2001. Some cladistic also work prefer Anthracosauria. (Gauthier et al 1988)
In 1956 Friedrich von Huene included both amphibians and anapsid reptiles in the Reptiliomorpha. This included the following orders: 1. Anthracosauria, 2. Seymouriamorpha, 3. Microsauria, 4. Diadectomorpha, 5. Procolophonia, 6. Pareiasauria, 7. Captorhinidia, 8. Testudinata. (Von Huene 1956) Now we know that these groups belong to very different lineages, and similarities are the result of convergences or shared primitive features.
In 1997 Michel Laurin and Robert Reisz (1997) adapted the term in a cladistic sense. Michael Benton (2000, 2005) made it the sister-clade to Batrachomorpha. However, when considered a linnaean ranking, Reptiliomorpha is given the rank of superorder and only includes reptile-like amphibians, not their amniote descendants (Systema Naturae 2000). More recently Reptiliomorpha has been adopted as the term for the largest clade that includes – according to the technical definitions of the phylocode which only refers to species level organisms – Homo sapiens but not Ascaphus truei (a primitive frog) (International Phylogenetic Nomenclature Meeting 2003); or is, as Palaeos co-author Toby White puts it, more like dogs than frogs (i.e. mammals but not amphibians). However, given the lack of consensus of the phylogeny of the labyrinthodonts in general, and the origin of modern amphibians in particular, the actual content of the Reptiliomorpha under the latter definition is uncertain. MAK050702 and updated by editors on Wikipedia, MAK111107
Reptiliomorpha: dogs > frogs.
Range: from the Early Carboniferous.
Tetrapoda*: Lepospondyli + * : Anthracosauroidea + (Gephyrostegidae + Batrachosauria)
Domed skull, retaining kinesis (skull roof loosely attached to cheek); skull usually with fine radiating grooves; tabular large & attached to parietal; deep otic notch in quadrate; some with slight, impedance-matching stapes; pleurocentrum dominant element of vertebrae; well-developed limbs; 5 digits; usually terrestrial.
Image: Silvanerpeton miripedes Clack 1994 Visean of Scotland, about 40 cm long, from David Peters, Reptile evolution, references Clack 1994, Ruta & Clack2006 MAK111107
Links: lecture9.
Note: A reminder that we are referring to the crown group Tetrapoda*, i.e. frogs + dogs. Reptiliomorpha is one stem from the tetrapod node by definition. The other stem really ought to be called something other than Lepospondyli, since we really don't know if the traditional lepospondyls included the ancestor of modern amphibians. ATW021214.

Anthracosauroidea: Eldeceeon, Silvanerpeton
Reptiliomorpha : (Gephyrostegidae + Batrachosauria) + * : Eoherpeton + (Chroniosuchia + Embolomeri).
Characters: "Kinetic line," i.e. abrupt angle between skull table and cheek without interdigitating suture & possibly mobile (primitive) [C02]; tabular contacts parietal [C02]; tabular horn or process common [C02]; intertemporal present (primitive) [C02]; "otic notch" present; posttemporal fossae absent; stapes massive [C02]; still has sarcopterygian skull hinge; moveable joint at basicranial articulation primitive) [C02]; closed palate without vacuities (primitive) [C02]; vertebrae gastrocentrous, with pleurocentrum dominant element [C02]; lozenge-shaped interclavicle; 5-toed hand [C02]; ilium with 2 dorsal blades [C02]; amphibian ankle; 2-3-4-5-3 almost reptilian) phalangeal count.
Note: In the figure, the intercentrum is shaded. The rachitomous type is dominated by the intercentrum, and the pleurocentra appear as paired, dorsolateral elements cushioning the notochord.. It is highly flexible and permits simultaneous twisting and bending -- appropriate for aquatic mobility, but likely to collapse on land. In the gastrocentrous vertebra, the emphasis is on supporting the arch. The neural arch sits firmly on the intercentrum and the pleurocentrum is reduced to a small, spacer element. This configuration allows less mobility, but assists the spine to bear the weight of the body on land. The embolomerous vertebra is characteristic of the secondarily aquatic embolomeres. The intercentrum reappears as a major structural unit, and both centra form complete disks. This conformation with successive small disks, recovers some of the flexibility of the rachitomous vertebra, while maintaining considerable strength and stability.
References: Clack (2002) [C02]. ATW020620.
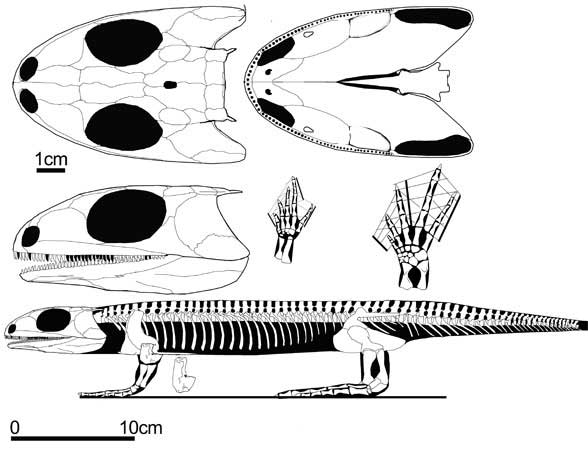
Eoherpeton: E. watsoni Panchen 19??
Range: Early Carboniferous (Namurian) of Europe (Scotland).
Phylogeny: Anthracosauroidea : Chroniosuchia + Embolomeri + *.
Characters: otic notch absent?
Image: (upper) Eoherpeton by Mike Coates; (lower) Eoherpeton skull, right lateral and dorsal view, from Smithson, TR (1985b), The morphology and relationships of the Carboniferous amphibian Eoherpeton watsoni Panchen. Zool. J. Linn. Soc. 85: 317-410; vide Clack (2002),
 Links: EMBOLOMERI; Crassigyrinus scoticus.
Links: EMBOLOMERI; Crassigyrinus scoticus.
Note: Eoherpeton has been variously classified as a gephyrostegid, an embolomere, and an anthracosaur i.s. It may also be none of the above. By way of explanation, this is one of these awkward areas in which we must recall our principles, even if we don't stand by on them. If Reptiliomorpha is defined as a stem group (dogs > frogs), then Anthracosauroidea must logically be a crown group (Gephyrostegus + Pholiderpeton). There is no guarantee that Eoherpeton falls in this group, particularly since the gephyrostegids are not very well known. It is included here among the anthracosauroids only by convention. ATW020621, MAK981010.
checked ATW040109