Aves: Neornithes
Taxa on This Page
- Lithornithiformes X
- Neornithes
- Paleognathae
- Tinamiformes
Neornithes: Running Out of Time
 A surprising number of recent papers in avian evolution still treat the Cretaceous evolution of birds as if it were a stately, linear progress from waddling dinosaurs to the near-miracles of neornithine flight engineering. Of course paleontologists know better. Evolution is never "toward" anything in a teleological sense. However, it is admittedly hard to keep this in mind at all times. Our every-day experience is of eagles and swifts and other birds so highly evolved for flight that it seems absurd to think they might ever have been specialized for anything else.
A surprising number of recent papers in avian evolution still treat the Cretaceous evolution of birds as if it were a stately, linear progress from waddling dinosaurs to the near-miracles of neornithine flight engineering. Of course paleontologists know better. Evolution is never "toward" anything in a teleological sense. However, it is admittedly hard to keep this in mind at all times. Our every-day experience is of eagles and swifts and other birds so highly evolved for flight that it seems absurd to think they might ever have been specialized for anything else.
However, just as an exercise, we started running (or flying, as the case may be) down the list of probable Cretaceous neornithines. The Cretaceous was full of flying birds, and also appears to have been full of various neornithine birds. The interesting point is that there were rather few flying neornithine birds from the Cretaceous. This requires a little qualification. The neornithines evolved from potentially volant avian stock. So it is likely that the earliest neornithines flew. For that reason we exclude the Lithornithiformes who seem to have been at or close to the very base of the neornithine radiation. However, it is worth noting that a remarkable proportion of other birds close to the base of Neornithes were non-flyers: e.g., Patagopteryx, Hesperornis, Otogornis (Hope, 2002).
But, after Lithornis and company, we have the unclassifiable mihirung (Dromornithifirmes) and the Phorusrhachidae, both of which probably had Cretaceous roots. Both could probably fly as well as a helium-filled hippopotamus. Then we might list the tinamous (Tinamiformes), which can become airborne in emergencies, but are famous for flying full-tilt into trees and breaking their necks when they do so. Then all the rest of the ratites, none of whom can fly any better than we can. But how about the neognaths? The Cretaceous probably had some early galliforms. Chickens can fly a bit, usually better than the tinamous; but many turkeys can't even manage this. Megapodes and the like are accomplished runners, but also weak flyers. On the anseriform side, we do better [1]. There are still a number of flightless forms, but most ducks and geese are passable flyers, although more highly adapted to water than air. Further, we can be certain that duck-like birds (Vegavis) were present in the very latest Cretaceous. Clarke et al. (2005). However, until the very end of the Cretaceous, the number of fully volant neornithines seems remarkably small.
Contrast this spotty record with the main competitors of Neornithes: the Enantiornithes. The argument that enantiornithines were poor flyers is no longer tenable. Chiappe & Walker (2002). In fact, we can now stand the argument on its head. If, as seems to be the case, the basic enantiornithine body plan is volant, what is the evidence that any enantiornithine was flightless? As far as we are aware (a major reservation!) no such evidence exists. Thus, we cannot blame the fossil record for all those earth-bound neornithines. The overwhelming weight of the evidence -- and of the birds, for that matter -- is that the neornithines initially specialized in various niches which required little or no flight.
Finally, we might speculate that all this running around was the ultimate cause of neornithine success. As in acting, good bones and staying thin will only get you so far. To really make use of the talent, you have to have high energy. The neornithines kept close enough to flight [2] to retain the good bones and, (with some exceptions -- see image) they kept thin. What the running lifestyle may have given them was the more completely endothermic metabolism needed to make good use of the ability to fly.So, when neornithines returned to the air in the Latest Cretaceous, they were pre-adapted for a whole new range of high-energy flying behaviors not open to the Enantiornithes, such as migration, predation on the wing, or extended care of young. ATW051030
Speaking of running around, Tsuda et al. 2007) have recently reviewed and extended some work on the sex chromosomes of basal neornithines. The bird sex-determination system is the reverse of the mammalian system. Females are heterozygotic, having both Z and W chromosomes. Males are ZZ. As Tsuda et al. reconstruct the history, early neornithines had sex chromosomes (the W and Z chromosomes in birds) which were very similar, differing not much more than ordinary pairs of somatic chromosomes in size and gene content. This similarity is retained to some extent in tinamous and ratites. In both of these groups, some Z-specific ("male") genes are deleted on the W chromosome, the gaps being filled by repetitive, non-coding sequences. Oddly enough, the patern of deletion seems to begin near the middle of the chromosome and work outward over the course of bird evolution. In neognathous birds (all living birds except ratites and tinamous), the Z and W chromosomes are highly differentiated.
This finding tends, if slightly, to confirm our speculation above. That is, it provides another indication that the primitive condition for neornithines was similar to that of ratites and tinamous: with only slight sexual dimorphism and, so, perhaps also a primarilly ground-based, non-flying mode of life. ATW080115.
Notes: [1] We exclude the screamers. The Anhimidae are quite good flyers today, but have a number of characteristics so weird that we question whether their immediate Cretaceous ancestors could fly at all: lack of apteria, ground-nesting, large number of cervical vertebrae, hallux low, very large feet.
[2] Ostriches and a few other non-flying birds with substantial wings seem to spread their wings as each leg passes the peak of the swing phase at in the running cycle (pers. obs.). This makes good mechanical sense, since the air resistance of the wings spread horizontally (a) reduces the ground reaction force (b) increases the time in the air, (c) may contribute to balance and maneuverabiliity, and (d) allows the the bird to remain more stiff-legged which, in turn, improves mechanical advantage (Hutchinson, 2004). At some point, wing reduction may become so pronounced that this sort of effect is no longer significant. However, until then, the interplay between limb and wing in fast locomotion may lead back to flight.
 Neornithes: modern birds.
Neornithes: modern birds.
The more important anatomical characteristics of this group are discussed in the Overview. They include: horn beak; teeth absent; fused limb bones. In addition Neornithes have a fully-separated four-chambered heart and typically exhibit complex social behaviors.
Links: Systematics of the Aves; Neornithes; Field Guide to the Birds on the Web (of the many lists of bird links on the Web, this seems to have more of scientific interest); Fish and Wildlife Information Exchange--Species Information; ORNICOLTURA ITALIANA; BIRDS TROUGHOUT TIME; Birding - Links - Suite101.com; BIRDNET: all about Ornithology, the Scientific Study of Birds; BIRDS; Neornithes - orders; Neornithes; Neornithes - Biognomen; Systematics of the Aves; Birds.
Image: © 1999 BIODIDAC and reproduced per standard terms of use. ATW020919
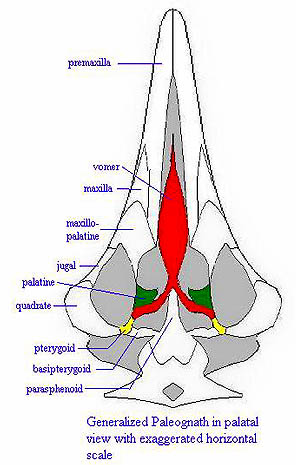 Paleognathae: Ratites and their ancestors.
Paleognathae: Ratites and their ancestors.
Range: From the Paleocene.
Characters: $ "Paleognathous" palate, consisting of: large pre-?)vomer attached to pterygoid and palatines, excluding both from parasphenoid joint between , no pterygoid and palatine; moveable joint between braincase and pterygoid, and large basipterygoid processes; primitively, splenial present on dentary; numerous "flight" characters of skeletal anatomy are generally preserved, if more or less vestigialized. May be polyphyletic -- various phylogenetic possibilities discussed above. Most relevant characters are likely primitive for neornithines. For example, the paleognathous palate is known from Hesperornis. Thus, there are no definite synapomorphies of this group. The figure at right is loosely based on a stylized rhea.
Links: Struthio mtDNA sequence; 75th Abstracts -- Abstract 34.
Lithornithiformes:
Lithornis, Pseudocrypturus.
Range: Early Paleocene to Middle Eocene of North America & Europe.
Early paleognaths, generally very similar to tinamous but clearly flying, perching forms. Splenial present; rhynchokinetic skull; cranial bones relatively unfused; carinate sternum; tail small with weakly fused pygostyle; large, curved claws.
 Tinamiformes:
Eudromia, Crypturellus.
Tinamiformes:
Eudromia, Crypturellus.
Living quail-like paleognathous flyers. Small head; rhynkokinetic skull; paleognathous palate; medium bill; long neck; carinate, with sternum similar to galliforms; heavy body with very large, abundant rump feathers; tail small; short, round wings; legs thick; claws flat; 3 toes, with 4th raised or absent; heart and lungs small for size; bill & leg colors often vivid and diagnostic; fast runners but easily exhausted; weak flyers (up to 100m) but navigation poor; diet varies, including seeds, roots, insects, berries, frogs and mice; frequently dig for food with bill, not legs; usually simple vocal behavior; inhabit all inland terrains including jungle and mountains to 4000m; presently known from Mexico to Argentina; male builds and guards nest, incubates eggs.
.Image from Knowlton (1909), with thanks to Robert. B. Hole.
Note: It is said to be dangerous (to the bird) to startle tinamous in forested areas, as they may fly into trees like an avian George of the Jungle. ATW011203.