The Cretaceous Period - 1
The Cretaceous Period of the Mesozoic Era: 146 to 65.5 million years ago
In 1822 the Belgian geologist D'Omalius d'Halloy gave the name Terrain Cretace, for the chalk and rock outcrops of the Paris Basin, and for similar deposits in Belgium and Holland and traceable also from England eastward into Sweden and Poland.
This term "Cretaceous" "chalk-bearing" (from Creta, the Latin word for chalk) later came to be used. The famous White Cliffs of Dover, are typical of this rock formation. Extensive chalk deposits were laid down in Europe and parts of North America during this period. The chalk itself is actually formed from the shells of countless micro-organisms.
William Smith had previously mapped four strata between the lower clay (= early Tertiary) and the "Portland Stone" (= late Jurassic), namely White Chalk, Brown or Grey Chalk, Greensand, and Micaceous Clay or brick earth (later referred to as Blue Marl or Gault). In 1822 Conybeare and Phillips arranged these in two groups, the Chalk and the earlier strata, a division that has remained to the present day. In 1841 Leymerie introduced the term Neocomian for the lower division. Senonian was coined by d'Orbigny in 1842 for the later Cretaceous. The name "Gallic" has also been used for times that do not fall conveniently into either of the above two categories.
Much of the land masses are covered by shallow continental oceans and inland seas. Europe, Asia, Africa and North America are all a series of islands. Marked regional differences in floras and faunas between the northern and southern continents developed, and indeed there were even differences in flora and fauna from continent to continent.
The breakup of the world-continent Pangaea, which began during the Jurassic, continued apace.
The Cretaceous saw the lengthening and widening of Atlantic Ocean, which especially spread further south, while the Alps were formed in Europe. India broke free of Gondwanaland and became an island continent, much like Australia is now. Africa and South America split apart, Africa moving north and closing the gap that was once the Tethys sea. The continents began to take on their modern forms.
| Period |
Epoch (ICS, with
added subdivision) |
Harland Epoch |
Age |
ICS Base |
ICS Duration |
| Paleogene |
Paleocene:
9.7 My |
Paleocene |
Danian |
65.5 |
3.8 |
Cretaceous
80.0 My |
Late Cretaceous II
End-Cretaceous)
18.0 My
|
Senonian |
Maastrichtian |
70.6 |
5.1 |
| Campanian |
83.5 |
12.9 |
Late Cretaceous I
The "High Cretaceous")
16.1 My
|
Santonian |
85.8 |
2.3 |
| Coniacian |
89.3 |
3.5 |
| Gallic |
Turonian |
93.5 |
4.2 |
| Cenomanian |
99.6
|
6.1 |
Early Cretaceous II
Aptian-Albian)
25.4 My
|
Albian |
112.0 |
12.4 |
| Aptian |
125.0
|
13.0 |
Early Cretaceous I
"Neocomian")
20.5 My
|
Barremian |
130.0
|
5.0 |
| Neocomian |
Hauterivian |
136.4 |
6.4 |
| Valanginian |
140.2 |
3.8 |
| Berriasian |
145.5 |
5.3 |
| Jurassic |
Late Jurassic:
15.7 My |
Malm |
Tithonian |
150.8 |
5.3 |
This site originally used the Harland three-fold division of the Cretaceous into Neocomian, Gallic, and Senonian Epochs. As of this writing (040911) we are in the process of gradually converting to the ICS system, which recognizes only Early and Late Cretaceous epochs. Unfortunately, the ICS Cretaceous epochs are unreasonably long for our purposes, so we have taken the further step of dividing both of them into two.
Our "Neocomian" division is based largely on climatic considerations. This "Early Early Cretaceous" demi-epoch was a time of steadily rising seas and temperatures. According to one source, the ocean temperature increased an almost unbelievable 17 C° over the Early Cretaceous, and the bulk of this must increase must have occurred in the Neocomian division. The Aptian-Albian division continued the Neocomian trend, but at a slower rate. The Aptian-Albian is also the interval that produced the definitive Cretaceous dinosaur clades. These dinosaurs dominated the large herbivore guild in the Late Cretaceous: the ornithischian iguanodonts including hadrosaurs), Ceratopsia (e.g. Triceratops), and various saurischian titanosaurs. In the oceans, a final radiation of pliosaurs also occurred at about this time.
The Early Late Cretaceous ("High Cretaceous") was marked by several critical events. The first was the widening Atlantic rift. The Atlantic Ocean: (a) had become wide enough to become a complete barrier to east-west dispersal over its entire length, except in the far north, and (b) was circulating meaningful amounts of ocean water north and south. The initial results seem somewhat paradoxical. On the one hand, the High Cretaceous experienced unprecedented uniformity of ocean temperatures from pole to pole, suggesting very good horizontal mixing of ocean waters. On the other hand, it is well known for sporadic deep ocean anoxia, which would indicate poor vertical mixing. It is tempting to speculate on the causes of this peculiar set of events. One strong line of evidence implicates methane and/or carbon dioxide outgassing. But most of this data comes from the Atlantic basin. In due course, we will have enough information from the Pacific to give us a better global perspective.
The second major event of the High Cretaceous was angiosperm dominance. Angiosperm plants had begun to spread at least as early as the middle Neocomian. However, during the High Cretaceous, angiosperms reached some critical mass or critical stage of development and became the dominant type of plant in most parts of the world. Finally, the long, gradual increase in sea levels which began in the Triassic reached its peak in the High Cretaceous.
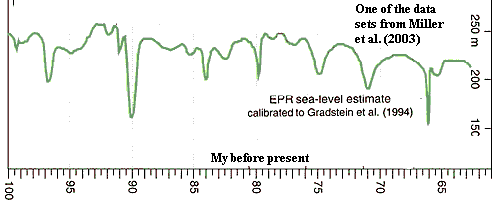 During the End-Cretaceous, sea levels began to retreat after 165 million years of advances. Miller et al. (2003) have recently reported that the peak and decline of sea levels in the Late Cretaceous is punctuated by a number of sudden, drastic, marine regressions. Apparently, the oceans retreated quite quickly, and, almost as quickly, returned to more or less their former depths. The pace of these changes appears to have been below the limit of geological resolution for their core samples, i.e. about <500Ky. Miller et al. state that their data are consistent only with the formation of short-lived, but rather extensive ice sheets in Antarctica. This conclusion is almost -- but not quite -- irreconcilable with what we know about Late Cretaceous climate. Miller's group coordinates data from a number of different regions in arriving at this result. While these sequences are diverse, they still cover only the Atlantic and Tethyan regions. Once again, we are badly in need of data from the Pacific Basin.
During the End-Cretaceous, sea levels began to retreat after 165 million years of advances. Miller et al. (2003) have recently reported that the peak and decline of sea levels in the Late Cretaceous is punctuated by a number of sudden, drastic, marine regressions. Apparently, the oceans retreated quite quickly, and, almost as quickly, returned to more or less their former depths. The pace of these changes appears to have been below the limit of geological resolution for their core samples, i.e. about <500Ky. Miller et al. state that their data are consistent only with the formation of short-lived, but rather extensive ice sheets in Antarctica. This conclusion is almost -- but not quite -- irreconcilable with what we know about Late Cretaceous climate. Miller's group coordinates data from a number of different regions in arriving at this result. While these sequences are diverse, they still cover only the Atlantic and Tethyan regions. Once again, we are badly in need of data from the Pacific Basin.
ATW040911, last revised ATW041024. Public Domain. No rights reserved.
Cretaceous Life
Small Stuff: Plankton &Microorganisms
 The Cretaceous was as long and eventful for microorganisms as it was for nearly everything else. For terrestrial macro-organisms, the critical factor may have been the gradual spread and eventual domination of angiosperm plants in the Late Cretaceous. For marine macro-organisms, the driver may have been the gradual rise of sea levels over the course of the Early Cretaceous. We are pleased to report that this happy karmic balance is retained in microorganisms, which experienced great changes related to various events (not all of which are understood) over the middle Cretaceous, specifically the Aptian-Albian episode and the earlier part of the High Cretaceous.
The Cretaceous was as long and eventful for microorganisms as it was for nearly everything else. For terrestrial macro-organisms, the critical factor may have been the gradual spread and eventual domination of angiosperm plants in the Late Cretaceous. For marine macro-organisms, the driver may have been the gradual rise of sea levels over the course of the Early Cretaceous. We are pleased to report that this happy karmic balance is retained in microorganisms, which experienced great changes related to various events (not all of which are understood) over the middle Cretaceous, specifically the Aptian-Albian episode and the earlier part of the High Cretaceous.
The main drivers at this time may have been changes in ocean chemistry, probably associated with the very active mid-ocean ridges of the time. Ocean crust was forming at a record rate, with all the usual geological belching and stretching that accompanies such events. Water temperatures became exceedingly, perhaps excessively, hot. Methane and carbon dioxide levels may have increased, and perhaps spiked at far above Early Mesozoic levels. Something seems to have happened to deep ocean circulation, since anoxic conditions prevailed at depth and black (poorly oxidized) shales formed from partially decomposed organic matter at several points during this middle part of the Cretaceous. Ocean fishes can often swim away from unpleasant environments. Single-celled organisms are bit more limited in their responses to deteriorating neighborhoods. Like reef biota or small grocery stores, they tend to adapt or die. Many died, but many adapted.
 One trend which is likely related to these events is the mineralization of many microorganisms. Whether unmineralized versions of these organisms were present before the mid-Cretaceous, is sometimes hard to tell. However, beginning in the Aptian-Albian, we see a sharp increase in the diversity and biomass of marine organisms with mineralized skeletons. This bears remembering because that is exactly what happened during the only other time in Phanerozoic history when the sea floor spread at this rate -- the Cambrian Explosion of Cambrian Epoch 2. At that time, the biological first response was the evolution of the "small shelly fauna" of small, mineralized creatures. At the same point in the Cretaceous episode, we find new foraminifera, new coccolithophores and other armored algae (e.g., silicoflagellates), and diatoms. We will briefly cover these forms and, finally, the Crenarcheota. This is a group so odd and so cryptic that their very existence was unknown until about 1998. Yet they are common in many oceans and may have played a major role in the events of the mid-Cretaceous.
One trend which is likely related to these events is the mineralization of many microorganisms. Whether unmineralized versions of these organisms were present before the mid-Cretaceous, is sometimes hard to tell. However, beginning in the Aptian-Albian, we see a sharp increase in the diversity and biomass of marine organisms with mineralized skeletons. This bears remembering because that is exactly what happened during the only other time in Phanerozoic history when the sea floor spread at this rate -- the Cambrian Explosion of Cambrian Epoch 2. At that time, the biological first response was the evolution of the "small shelly fauna" of small, mineralized creatures. At the same point in the Cretaceous episode, we find new foraminifera, new coccolithophores and other armored algae (e.g., silicoflagellates), and diatoms. We will briefly cover these forms and, finally, the Crenarcheota. This is a group so odd and so cryptic that their very existence was unknown until about 1998. Yet they are common in many oceans and may have played a major role in the events of the mid-Cretaceous.
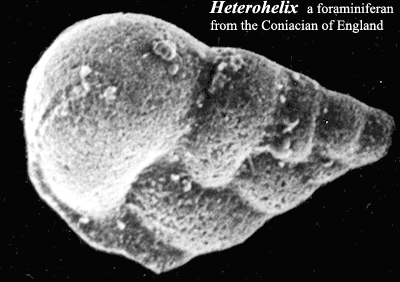
Foraminifera: of all the groups we will discuss, the forams were the least changed by the events of the Cretaceous. These are amoeboid protists with which evolved in the Early Cambrian -- possibly as part of the original small shelly fauna. They extend pseudopods through holes in their shell or, more properly test), to feed on bacteria and small food particles, like radiolarians. See, e.g., Acantharea. Also like radiolarians, they frequently live in symbiosis with photosynthetic protists, typically dinoflagellates. In the earliest forms, the test is formed simply by gluing together foreign particles. Simple shells appear in the Devonian, and the group  gradually became planktonic, rather than benthic, over the course of the Mesozoic. A major turnover of planktonic foraminifera occurred, associated with many extinctions, during the first Ocean Anoxic Event ("OAE") of the mid-Aptian. The net result was a great diversification of planktonic forms with elaborate calcareous tests. See, e.g., the image of Heterohelix.
gradually became planktonic, rather than benthic, over the course of the Mesozoic. A major turnover of planktonic foraminifera occurred, associated with many extinctions, during the first Ocean Anoxic Event ("OAE") of the mid-Aptian. The net result was a great diversification of planktonic forms with elaborate calcareous tests. See, e.g., the image of Heterohelix.
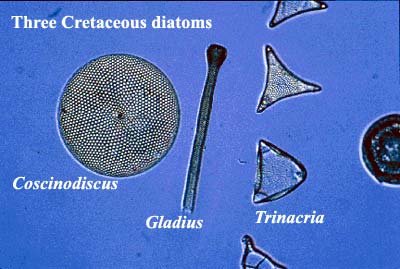
Diatoms: Diatoms are eukaryotic, photosynthetic microorganisms that, today, are found in both fresh and salt water bodies throughout the world. They are said to account for the majority of marine primary productivity, and as much as 20% of the total photosynthetic activity on Earth. Yet they did not exist in any numbers prior to the mid-Cretaceous.
Calcareous nanoplankton: The Cretaceous Period was named for the great Late Cretaceous chalks of Northern Europe. These were formed from the tests of calcareous nanoplankton (armored algae), of which the best known are the coccolithophores. Nanoplankton with mineralized "armor" are known from the Late Triassic or Early Jurassic. Their fortunes seem to be tied rather directly to sea levels, so that their abundance peaked in the Late Cretaceous -- perhaps a bit later than the Aptian-Albian events we are discussing -- but perhaps not. It is not possible to make quantitative comparisons with much precision. Coccolithophores are responsible for some algal blooms today. One trigger for their rapid growth appears to be dissolved iron, which is often a limiting nutrient for algal growth. One school 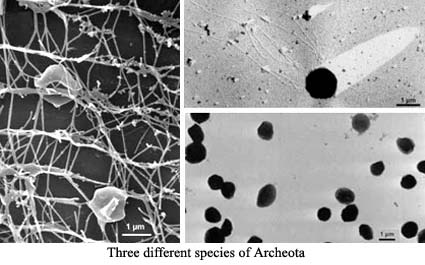 holds that iron released by new continental crust may have been responsible for massive blooms leading to the Mid-Cretaceous black shales and anoxic deep-ocean conditions. At any rate, some of the recoverable organic compounds from the black shales are biomarkers for haptophyte algae, a group that includes the coccolithophores. Bralower et al. (2002).
holds that iron released by new continental crust may have been responsible for massive blooms leading to the Mid-Cretaceous black shales and anoxic deep-ocean conditions. At any rate, some of the recoverable organic compounds from the black shales are biomarkers for haptophyte algae, a group that includes the coccolithophores. Bralower et al. (2002).
Why? The evolutionary "why" is a relatively easy question to answer. Tests provide some degree of physical protection, can be used to keep the cell in easy proximity to symbiotes, provide an ion reserve, can be used in at least two ways to control buoyancy, and allow extracellular storage of useful materials, among other advantages. The more difficult issue is why this became possible in the mid-Cretaceous and not before. What ocean chemistry made it easy to adopt such a difficult -- but ultimately useful -- piece of baggage?
Unfortunately, the answers involve rather complex geochemistry, which we are in no position to evaluate. One factor may have been the very low ratio of magnesium to calcium in Cretaceous seawater. This is another characteristic of the mid-Cretaceous which also applies to the Early Cambrian and was caused by the tendency of new ocean crust to lock up magnesium and release calcium. Calcite is nicely stable. Even an inefficient evolving) test producer can work with calcite in a high-calcium sea, since 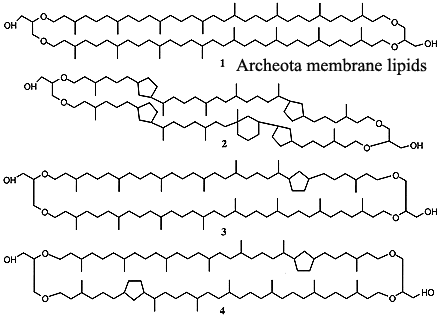 calcite will hold together and doesn't require much maintenance.
calcite will hold together and doesn't require much maintenance.
The Mysterious Archeota: One recent hypothesis explains some (not all) of the weirdness of the Mid-Cretaceous with even greater weirdness -- The Archeota. The Archeota are one of the two principle divisions of the Archaea. The Archaea, in turn, are the sister group of the Eukarya. The Archeota were discovered in the 1990's. They are quite small, and tend to live in slightly less bizarre environments than other Archaea. In particular, they tend to be thermophiles, but do not require anything like the hellish conditions under which other Archaea live. They cannot be cultured on conventional media. However, some can now be raised on a substrate of living sponges from which Archeote cells can be recovered in sufficient numbers and purity to do serious biochemistry. Although still poorly known, Archaeotes appear to be ubiquitous in the oceans. Schouten et al. (2000).
The critical finding is this: up to 80% of the recoverable organics from the black shales appear to be breakdown products from the unique membrane lipids of the Archeota. Any biochemist will assure you that these are very peculiar molecules. Unfortunately, the black shale organics are not these complete molecules, but even the breakdown products of these odd, cyclic diglycerol dialkylethers with inserted cyclopentane rings, are quite distinctive. It is not completely clear to us that these are precisely the same products described by Kenig et al. (2003) from both Archean (c. 2000 Mya) and modern sediments, but they seem closely related.
To add one more ingredient to the mix, it has recently been reported that archeotes mediate marine methane metabolism. Schouten et al. (2003). Specifically, archeotes work in tandem with sulfate- reducing bacteria to produce carbon dioxide and hydrogen sulfide from methane and sulfates under anaerobic conditions. The Mid-Aptian OAE, at least, is associated with a massive release of methane and an apparently large spike in world temperatures. This methane was, presumably, the abiotic product of processes in the mantle and was released during the frenzy of mid- ocean ridge spreading which occurred at this time. It is thus reasonable to speculate that the release of methane and associated temperature increase of the Mid-Aptian acted to trigger massive blooms of some relatively low temperature (for a thermophile) archeote and its sulfate reducing symbiotes. These microorganisms would eventually eliminate the excess methane, saving the Earth from long-term over-heating; but, in the process, would also generate enough toxic hydrogen sulfide to kill everything within a rather large radius of the archeote bloom.
It's a thought, anyway. But it still doesn't explain why everything developed silicate or calcite tests at this time.
Image Credits: Heterohelix from the MIRACLE site of University College, London. Cretaceous diatoms from the Cushman Foundation for Foraminiferal Research. Archeotes from Ciència Hoje. Archeote lipids from Damsté et al. (2002).
Links: Tectonics and plankton evolution during the Mid-Cretaceous; Foraminifera; CALCAREOUS MICROFOSSILS; Chapter 12, part 6, Lecture Notes, Life of the Mesozoic, Levin ...; Cretaceous nannoplankton evolution and diversity.
ATW041024. Edited ATW050526. Text public domain. No rights reserved.
Continued on Next Page