
Osteolepis. Artwork by Nobu Tamura, 2007. Wikipedia, GNU Free Documentation/Creative Commons Attribution license.
| Sarcopterygii | ||
| The Vertebrates | Osteolepiformes |
| Vertebrates Home | Vertebrate | Vertebrate |
Osteolepis. Artwork by Nobu Tamura, 2007. Wikipedia, GNU Free Documentation/Creative Commons Attribution license. |
One of the nice things that begins to happen at the level of the osteolepiforms is the standardization of the anterior bones in the snout. Primitive sarcopterygians, and particularly dipnomorphs, break this area up into a hopeless mosaic of tiny little pieces. However, as we proceed in the direction of the tetrapods, this area begins to look more and more "normal." Unfortunately, the steps in the transition were not clear initially, with the result that much of the older literature uses actinopterygian terminology which can't be reconciled with the fossil record as it is now known. In particular, the often-cited work of Jarvik on Eusthenopteron uses the older terminology. Jarvik refers to the major roofing bone between the orbits as the "frontal." In fact, it is clearly homologous to the tetrapod parietal. Essentially all current work uses the corrected terminology described by Schultze & Arsenault (1985). This is summarized in the following diagram.
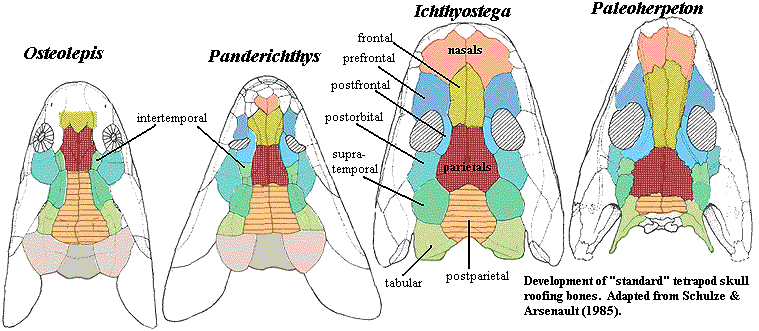
As mentioned in the discussion of Tetrapodomorpha, the development of a central arch of long bones from snout to occiput is one of the main themes of tetrapodomorph -- and tetrapod -- evolution. See, generally, Clack 2002). In the relatively basal osteolepidid Osteolepis, there is no definite skull table anterior to the parietal, which terminates between the orbits. The snout is short and broken up into a mosaic of small, irregular bones, rather like a lungfish. The parietal foramen is located between the orbits, on the anterior half of the parietals. The parietals meet the postparietals along a horizontal, unsutured line to accommodate flexing of the intracranial joint. The postparietals are long, but thin and loosely overlapping. The "transitional" bones, the intertemporals, supratemporals and tabulars, curve smoothly onto the plane of the cheek. The pre- and postfrontals are undistinguished and variable supraorbital bones, and the orbits are generally lateral.
In tetrapods, all this is changed. As the diagram from Schultze & Arsenault (1985) shows, the posterior skull -- the old otoccipital skull -- has been only slightly compressed. The loss of the extrascapular series, incident to the separation of the forelimb girdle from the skull, makes the posterior skull seem shorter. In fact, the principle change has been a lengthening of the ethmosphenoid region -- the anterior skull. The skull table has been extended by large, stout paired nasals and frontals. This arch from rostrum to occiput provides the basis for the enormous elongation of the rostrum, which is covered by a smaller number of regular paired bones. The parietal foramen is located well posterior to the orbits, which now face dorsally. The parietals have a complex, immovable suture with the postparietals, which are now small relative to the overall length of the skull. The transitional bones develop complex articulations with the skull table, which becomes wide and relatively flat. They maintain only a loose connection with the cheek, which angles sharply and abruptly downwards from the top of the skull. In many cases, the transitional bones and the cheek are separated by a deep extension of the old spiracular slit, referred to in tetrapods as the "otic notch." The pre- and postfrontals no longer look like extensions of the orbital series, but have taken on an independent role.
The sides of the skull, the "cheeks," also undergo considerable change. However, most of this is connected with the loss of the opercular series incident to the absence of gills in adult tetrapods. Since we are primarily concerned here with the non-tetrapod "fishy" forms, we will focus largely on the skull table.
These changes appear to have been initiated deep within the tetrapodomorph group, with a tendency to parallel development in the three main tetrapodomorph lineages. The rhizodonts are poorly known, but Barameda shows some evident movement in this direction. The skull of Barameda is loosely constructed, with a deep and definite break at the intracranial joint. However, the orbits are at the anterior end of the parietals, and two bones that may be definitive frontals are prominent features of a slightly elongate rostrum. The transitional bones suggest the absence of a sharp boundary between roof and cheek, but the postparietals have the beginnings of a complex suture between them, solidifying the posterior skull roof. The pre- and postfrontals are rounded and do not resemble the orbital series.
The tristichopterids have a far better fossil record, and it reveals some of the complexity of the selective forces at work. The skull of Eusthenopteron is little different from Osteolepis. In some ways, it seems more primitive. However, the anterior skull is distinctly elongated, an effect achieved by a uniquely large and broadened parietal and an equally unique large median rostral surrounded by an arch of smaller rostral bones -- apparently an autapomorphic tristichopterid solution to the general tetrapodomorph problem. The same general approach is seen in the derived tristichopterids, Mandageria and Cabonnichthys. Cabonnichthys in particular takes the tristichopterid "solution" one step further by superficial fusion (sutures are still observable internally) between the massive parietal and the small bones of the rostrum. Unlike the rhizodonts, the tristichopterids have a marked, open spiracular slit which creates a natural break between skull table and cheek.
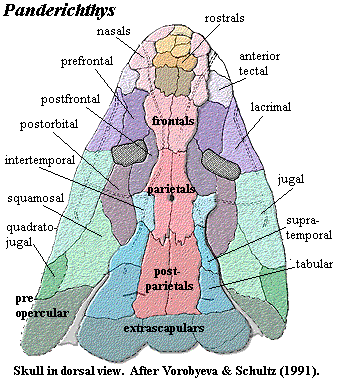
Finally, the Elpistostegalia show definitive paired frontals and a series of bones that appear to be early homologues of the nasals. See Panderichthys skull image. The spiracular opening is marked. Externally, the intracranial joint has been obliterated by a tight suture between parietals and postparietals. Here we also begin to see the tetrapod pattern on the lateral skull, with a significant reduction in the opercular series and the jugal at last meeting the quadratojugal, breaking the old contact between squamosal and maxilla. Perhaps even more significant than the pattern of skull bones is the manner in which they are assembled. Clack 2002). Rather than a relatively loosely assembled aggregation of overlapping plates, elpistostegids show many of the sorts of articulations we are accustomed to seeing in tetrapods: tight, interdigitating sutures, scarf joints, tongue-in-groove articulations and so on.
In general, the dermal bones of the skull show a gradual and concerted change over the course of phylogeny. This pattern is somewhat independent of lineage and thus probably reflects a true concerted, adaptive change applicable to a wide range of animals. We will not speculate -- at least not yet -- on what it might mean. ATW031029.
 |
|
Eusthenopteron foordi. Late Devonian (Frasnian), Escuminac Formation, Quebec, Canada (Western Laurasia). Length 1 metre. |
Its difficult to say what characteristics Osteolepiformes and Osteolepididae have because their definitions and positions have changed so often over the years. With respect to Osteolepiformes, we're more or less back to where we were a century ago, when Osteolepiformes was considered to be the order of Sarcopterygii ancestral to man. Translated into cladistic terms, that would be something like Rhinoceros > rhizodonts, which is the way the term is used here. Osteolepiforms are also frequently referred to as the Choanata, an apomorphy-based definition referring to the fact that the posterior nares has moved onto the palate as a choana. However, it is not certain that the Rhizodontiformes, or even the Porolepiformes, all lacked one, that the osteolepiforms all had one, or that the choana really is the homologue of the posterior nares. They are also called the Osteolepidida, a term perhaps intended to distinguish some crown group.
The Osteolepididae were once almost identical with the Osteolepiformes. Here, we have tried -- without notable success -- to restrict "Osteolepididae" to a small, probably monophyletic, group of fishes closely related to Osteolepis. Unfortunately, because of all this historical uncertainty about definitions, it is frequently unclear exactly what group is being discussed in the literature. Ultimately, we take the path of cowardice and convenience, treating both together as a paraphyletic grade of basal, non-rhizodont tetrapodomorphs. We will refer to the group generally as "osteolepids," a suitably vague and ambiguous term. For the following description it may be useful to summon up our usual cranial image of Eusthenopteron.
 The osteolepids all have some kind of frontal bone, basally not very well differentiated. The frontal is usually paired, but sometimes not, as in Eusthenopteron. Frequently, the frontals are accompanied by a series of one to four paired rostral bones which are said to be early homologues of the nasals. The nares are close to the margin of the jaw, but a small lateral rostral separates them from the edge. The dorsal side of the nares is formed by the anterior tectal. The terminology varies, but the lateral tectal, anterior supraorbital, and prefrontal appear to be all the same, and this lies posterodorsal to the anterior tectal. Below the prefrontal is a lacrimal, which is relatively large and distinct. The jugal and postorbital are of comparable size, but frequently short, so that the anterior cheek is dominated by these three block-like bones. The squamosal is very large and extends from the skull roof down to contact both the maxilla and quadratojugal. The dorsal margin of the squamosal meets, but does not suture with, the skull table, which primitively includes a well-developed gap to accommodate motion at the intracranial joint. The interopercular is not small but is invariably narrow and vertically oriented. The opercular and a subopercular of variable size bridge the gap between the cheek and the cleithral bones of the dermal pectoral girdle.
The osteolepids all have some kind of frontal bone, basally not very well differentiated. The frontal is usually paired, but sometimes not, as in Eusthenopteron. Frequently, the frontals are accompanied by a series of one to four paired rostral bones which are said to be early homologues of the nasals. The nares are close to the margin of the jaw, but a small lateral rostral separates them from the edge. The dorsal side of the nares is formed by the anterior tectal. The terminology varies, but the lateral tectal, anterior supraorbital, and prefrontal appear to be all the same, and this lies posterodorsal to the anterior tectal. Below the prefrontal is a lacrimal, which is relatively large and distinct. The jugal and postorbital are of comparable size, but frequently short, so that the anterior cheek is dominated by these three block-like bones. The squamosal is very large and extends from the skull roof down to contact both the maxilla and quadratojugal. The dorsal margin of the squamosal meets, but does not suture with, the skull table, which primitively includes a well-developed gap to accommodate motion at the intracranial joint. The interopercular is not small but is invariably narrow and vertically oriented. The opercular and a subopercular of variable size bridge the gap between the cheek and the cleithral bones of the dermal pectoral girdle.
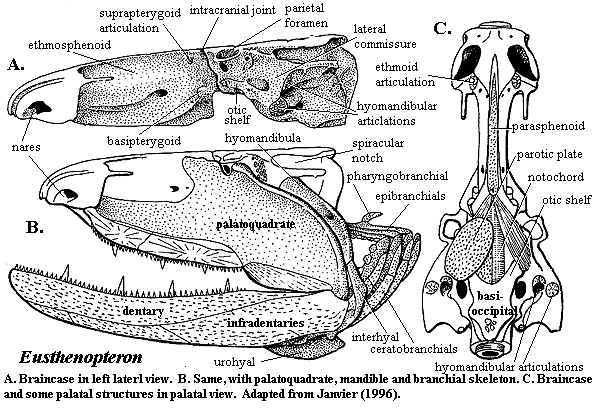
The osteolepid braincase is well-known. The braincase is cleanly divided into two segments. A remarkable feature of the braincase is the absence of bone ventrally in the posterior, otoccipital segment. The basisphenoid is virtually absent and the notochord is exposed both ventrally (in the anterior half of the otoccipital region) and laterally in the posterior half). In life, the anterior median vacuity was covered by dermal tooth plates, the parotic or arcual plates. The lateral vacuities are referred to as the vestibular fontanelles and may be the homologues of the middle ear. These spaces may have accommodated large muscles which flexed the intracranial joint, as shown in Janvier's (1996) reconstruction below. They communicated with the branchial region and body chamber through deep metotic fissures laterally, between the otic capsule and the body of the braincase. Anteriorly, the otic capsules joined with the braincase through a broad, superficial lateral commisure. The principal structural role of this structure seems to have been to support two articulations for the double-headed hyomandibular posteriorly.
The anterior, or ethmosphenoidal, portion of the braincase was narrow, supporting a primitively narrow denticulated parasphenoid. However, the anterior part of the region broadened out into two well-separated and well-ossified nasal capsules which also provided mechanical support for the rostrum and anterior palate. The palatoquadrate was tightly bound to the braincase through no less than four articulations: ethmoid, basipterygoid, suprapterygoid, and paratemporal (= otic shelf). The dentition included large fangs but the anterior fangs were primitively absent, and vomer lacked a strong posterior process. The basibranchium included two stout elements and was prolonged anteriorly as a sublingual rod.
We will defer discussion of the axial and appendicular skeleton. The former was quite variable among osteolepiforms groups, and the latter is not well known in basal osteolepids. Primitively, osteolepids retained a thick layer of cosmine over the scales and exposed bone. However, the sensory pores were much less well developed than in porolepiforms, and the enamel layer did not extend far into the pores. In addition the osteolepids all possessed enlarged bony scutes at the bases of all paired and unpaired fins, a feature from which they derived their name. ATW031031.
 Osteolepiformes: (= Choanata = Osteolepidida) Gyroptychius, Medoevia.
Osteolepiformes: (= Choanata = Osteolepidida) Gyroptychius, Medoevia.
from the Middle Devonian (Eifelian) [CA96]. Possibly as early as Early Devonian (Emsian) [J80] [J96].
Tetrapodomorpha: Rhizodontiformes + *: Osteolepididae + (Megalichthyidae + (Canowindridae + (Tristichopteridae + Elpistostegalia))).
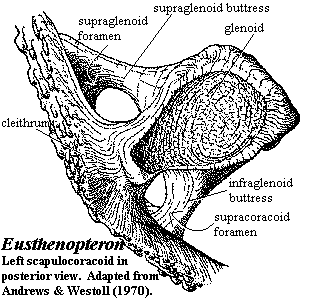
head large; nasal capsules with three exits (naris, choana, lacrimal duct) [C88] [VS91]; single external pair of nares [AM94]; nares with dermintermedial process [J96]; lateral rostral ventral to naris [VS91]; several narrow submandibulars [VS91]; lacrimal large [VS91]; lacrimal, jugal & postorbital of comparable size [VS91] (contra, [JA97]); large squamosal separates narrow preopercular from maxilla [CA96]; squamosal meets maxilla & separates jugal from quadratojugal [C02]; squamosal meets, but does not suture with skull table (supratemporal, tabular, postspiracular, etc.) [AJ97]; preopercular bar-like and almost vertically oriented [VS91] [JA97]; otico-occipital braincase retaining metotic fissures terminating in large vestibular fontanelle [CA96]; palatoquadrate articulating with braincase at ≥ 4 points: ethmoid, basipterygoid, suprapterygoid & paratemporal (= otic shelf) [J96]; spiracular canal lined with minute dermal bones [J96]; braincase divided by intracranial joint running through foramen for profundus branch of Vth nerve [CA96]; deep sella turcica [J96]; undivided anterior palatal recess [VS91]; choanae present, defined by vomer, palatine, maxilla & premaxilla [VS91] [CA96]; parasphenoid long and meets vomers [VS91] (contra, [JA97]); vomer posterior process present [CA96$] [3]; parasymphysial tooth plate [VS91]; large parotic plates present [J96]; coronoids often separated by deep fossae [AJ97]; prearticular exposed anteriorly to symphysis [A+00]; median gular present [VS91]; labyrinthine enamel; dentition primitively simple (not "labyrinthodont") [AW70a] (contra [C88]: basal rhipidistian character); anterior fangs primitively absent [AJ98]; five branchial arches [J96]; two basibranchial elements with underlying urohyal [J96]; basibranchium prolonged anteriorly as sublingual rod [J96]; body cylindrical; many had paired crescentic vertebral centra; dorsal ribs [VS91] [2]; vertebrae either rachitomous or ring-like [CA96]; ribs short or absent [CA96]; enlarged scutes at base of all fins except caudal [C88] [VS91] (but [CA96]: in the majority of taxa); first dorsal fin probably did not articular with spine & and is not well known in any genus [AW70a]; paired fins with short, uniserial, metapterygial skeleton [CA96]; fin radials unjointed & unbranched [CA96]; paired fins with preaxial radials only [CA96$]; three bones in extrascapular series, with lateral extrascapulars overlapping median [VS91]; well-developed anocleithrum [VS91]; anocleithrum exposed on surface [CA96$]; tripodal scapulocoracoid [J96] with large supraglenoid & suracoracoid foramina (see image) [J96]; scapulocoracoid dorsoventrally flattened with transversely elongate glenoid [J96]; pectoral fins strongly ossified [L95]; humerus with ball-in-socket articulation [VS91] [1]; primitively thick rhombic scales covered with cosmine, but lost in several lineages [AW70a] [L95] [CA96]; flask-shaped cavities in cosmine generally fewer and not lined with cosmine.
Notes: [1] As stated, this is clearly incorrect. What [VS91] may mean is that the humerus has a distinct convex head which fits into a glenoid fossa. [2] when ribs are present? [3] Cloutier & Ahlberg [CA96] state that the posterior process of the vomer may be absent in primitive osteolepidids and may represent a less-inclusive clade.
Links: Biology 356.
References: Ahlberg & Johanson (1997) [AJ97]; Ahlberg & Johanson (1998) [AJ98]; Ahlberg & Milner (1994) [AM94]; Ahlberg et al. (2000) [A+00], Andrews & Westoll (1970a) [AW70a]; Carroll 1988) [C88], Clack (2002) [C02], Cloutier & Ahlberg (1996) [CA96], Janvier (1980) [J80], Janvier (1996) [J96], Johanson & Ahlberg (1997) [JA97], Long (1995) [L95]; Vorobyeva & Schultze (1991) [VS91]. ATW031004.
checked ATW021227
{kind=link}
{kind=link}
{kind=link}