 Type C: (right side of Bob) the septomaxilla lies almost entirely external to the naris. It is sutured to the nasal and extends ventrally or laterally to partially obscure the naris.
Type C: (right side of Bob) the septomaxilla lies almost entirely external to the naris. It is sutured to the nasal and extends ventrally or laterally to partially obscure the naris.| Bones: Dermal Bones | ||
| Vertebrates | Facial Series: Septomaxilla-1 |
| s ("Cladograms") |
||||||
| Vertebrates Home | Vertebrate | Vertebrate | Bones | Time |
|
Facial Series
|
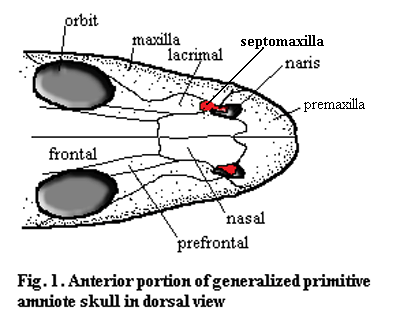
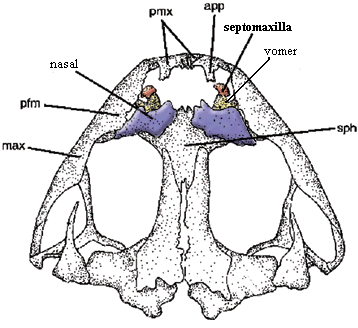
The septomaxilla is a small, usually delicate bone which is closely associated with the nares. In most cases some or all of the bone lies below the level of the dermal surface bones and inside the narial opening. On the right is the familiar dorsal shot of the snout of Bob, the Basal Amniote. In order to illustrate the obscure workings of the septomaxilla, we have been forced to magnify Bob a bit, which must surely gratify his basal amniotic ego. However, we have offset this minor flattery by making him asymmetrical. In fact, we have attempted -- albeit with limited success -- to illustrate three basic types of septomaxilla on one individual. With some exceptions, these three types are fairly distinct, and we will refer to them by letter designation.
Type A: (left side of Bob) the septomaxilla is often sutured to the lacrimal and appears to be an extension of that bone. Alternatively, it has a long external, posterior process towards the orbit. In either case, it is probably associated with the nasolacrimal duct. In this morph, it may or may not have additional structure inside the naris.
Type B: the septomaxilla is entirely inside the naris. It lies on the lateral or ventral inner surface, in contact with the underlying vomer. In this position it often bears a distinct dorsally or medially - directed process which, from the outside, appears to divide the naris into anterior and posterior compartments.
Type C: (right side of Bob) the septomaxilla lies almost entirely external to the naris. It is sutured to the nasal and extends ventrally or laterally to partially obscure the naris.
Perhaps the differences between these types may be explained by the different inductive influences these septomaxillae experience during development. The Type A bone is seems to function as an extension of the lacrimal and/or the nasolacrimal duct. Type B works with the vomer to form the floor of the naris and the shunt out of the nasal passage to the vomeronasal organ, or possibly the choana. Type C is a conventional dermal roofing bone which extends the nasal out over the naris. Thus, Types A, B, and C could be thought of as, respectively, nasolacrimal, vomeral and nasal septomaxillae. Our type designations have some phylogenetic utility, but they are not infallible. In some cases, as we will see, the type flips back and forth in a way which would be unbecoming of a reliable synapomorphy.
There are a number of bones which, by convention, are called "septomaxillae" but may well be neomorphs. For example, phytosaurs have a pair of small median bones, sutured on the midline. These are actually located anterior to the nares, which are dorsal and closely spaced. They are referred to as septomaxillae. In fact, they might actually be septomaxillae of a modified Type C, since they are flanked by the nasals. However, the phytosaur nares are unique, and it is difficult to be certain.
Finally, since septomaxillae are usually small, fragile bones, they are simply not recovered in many cases. In many forms, they may also 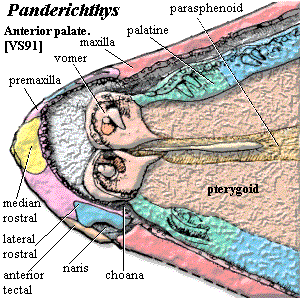 fail to ossify. As a result, the phylogenetic distribution of the bone is uncertain.
fail to ossify. As a result, the phylogenetic distribution of the bone is uncertain.
The three different structural morphs of the septomaxilla reflect its differing roles in various branches of the tetrapod tree. The Type A septomaxilla primarily involved in maintaining the nasolacrimal duct, either alone or in combination with the lacrimal. The Type B bone is similarly employed together with the vomer in reaching the vomeronasal organ and/or the choana. Both the B and C types -- perhaps all three types -- may also anchor muscles which close off or flare the nostrils. Clack (2002).
In the high and far-off days of the Middle Devonian the immediate ancestors of the Rhipidistia experienced some unknown selective force which caused the conventional pattern of rostral bones to fragment into a kaleidoscope of random-looking shards of bone. In the Dipnomorpha, the process eventually disassembled virtually all of the conventional dermal bones of the skull. Among our own ancestors, the Tetrapodomorpha, the process stopped early on, and eventually reversed course. In the transition back to a tidier skull, the tetrapodomorphs developed regular, if unconventional, sets of bones between the tip of the rostrum and the orbit -- series which were gradually simplified over the course of time and phylogeny. The two series which concern us are 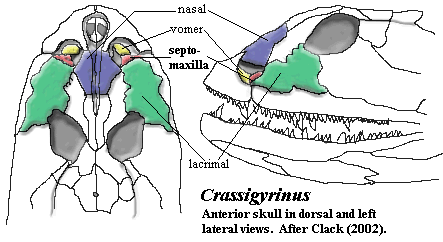 the rostral series, on and around the edge of the rostrum, and the tectal series, which occupied the more dorsal regions which would later be occupied by the nasal and lacrimal.
the rostral series, on and around the edge of the rostrum, and the tectal series, which occupied the more dorsal regions which would later be occupied by the nasal and lacrimal.
By the time we reach Panderichthys, on the borderlands of the Tetrapoda, four bones are in contact -- or near contact -- with the nares: the premaxilla, the maxilla, the lateral rostral, and the anterior tectal. One of the latter two bones is the immediate precursor of the septomaxilla. Clack (2002) (anterior tectal); Schultze & Arsenault (1985) (lateral rostral). The anterior tectal makes a more convincing precursor for a Type A or C septomaxilla, but the lateral rostral looks more like a Type B septomaxilla.
As shown in the image from Vorobyeva & Schultze (1991), the premaxilla had a tendency to slip in under the lateral rostral and meet the maxilla directly. All that would be required for a Type B septomaxilla is for the premaxilla to, instead, overlie the lateral rostral. Conversely, the anterior tectal is already behaving exactly like a Type C septomaxilla. So, since we lack any living osteolepiform fishes with which to experiment, it is impossible to say which is the more plausible candidate [1].
 Phylogeny of the Septomaxilla
Phylogeny of the SeptomaxillaThe first tetrapod septomaxillae which we meet are plainly of Type C, which favors a tectal origin. Both Acanthostega and Ichthyostega have this morphology. Clack (2002) [2]. However, Crassigyrinus, which may also be quite basal (or not!), has a very different septomaxilla, essentially of Type B. The septomaxilla lies entirely within the naris and below the surface of the dermal bones (id.), it bears a small dorsomedially directed process, and it is closely associated with the vomer. The two help form an unusual external (but probably subdermal) duct leading into the oral cavity. However, the septomaxilla is also in an essentially posterior position and flush with the lacrimal, as in Type A. Thus Crassigyrinus is somewhat anomalous, as it is in many other ways.
Among the baphetids, the septomaxilla is known in Megalocephalus. Here, it is essentially in the conventional Type B position. It sutures only (and narrowly) with the maxilla and premaxilla. Beaumont (1977). The bone appears depressed below the general surface of the skull in reconstruction. However, it is ornamented and thus presumably had some surface exposure. Thus, given the evidence from Crassigyrinus and Megalocephalus, one could question whether this is really the same bone as the septomaxilla of Ichthyostega. It might perhaps be a different ossification, derived from the lateral rostral.
 However, a brief review of the temnospondyls shows no pattern at all. Many temnospondyls either lack a septomaxilla, or the bone has never been recovered. It appears to be absent both in the basal Dendrerpeton and the derived Parotosuchids. Steyer (2002). When present, it is quite variable in size. Id. It may be entirely internal (Trematopidae) or have broad surface exposure (Thoosuchus). Clack (2002), Shishkin et al.
(2000). One rather revealing exercise is to compare Thoosuchus yakovlevi, as described by Ryabinin in 1925, with precisely the same specimen, as described by Shishkin et al. in 2000. The position and dermal exposure of the septomaxilla is entirely different in the two. The septomaxilla is, once again, a small and delicate bone and it is often only weakly sutured to other components of the skull. Thus, it is not always possible to assume that a particular reconstruction is accurate at the level we are examining this bone.
However, a brief review of the temnospondyls shows no pattern at all. Many temnospondyls either lack a septomaxilla, or the bone has never been recovered. It appears to be absent both in the basal Dendrerpeton and the derived Parotosuchids. Steyer (2002). When present, it is quite variable in size. Id. It may be entirely internal (Trematopidae) or have broad surface exposure (Thoosuchus). Clack (2002), Shishkin et al.
(2000). One rather revealing exercise is to compare Thoosuchus yakovlevi, as described by Ryabinin in 1925, with precisely the same specimen, as described by Shishkin et al. in 2000. The position and dermal exposure of the septomaxilla is entirely different in the two. The septomaxilla is, once again, a small and delicate bone and it is often only weakly sutured to other components of the skull. Thus, it is not always possible to assume that a particular reconstruction is accurate at the level we are examining this bone.
Living amphibians are. as usual, too derived to be of much help in working out early phylogeny. The frog septomaxilla appears to be of Type B, but the skull is so highly modified that this is not a particularly meaningful statement.
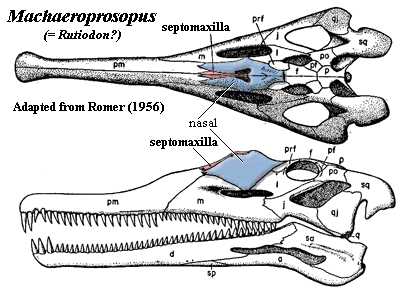
According to Romer (1956), the plesiomorphic condition for the amniote stem lineage is Type A. This is a bit peculiar, since this is the one type we have not seen much of in the basal tetrapods (Crassigyrinus being perhaps the only exception). However it makes good sense in a mechanistic kind of way. The Type A septomaxilla is involved in the nasolacrimal duct. The nasolacrimal duct is important only for truly terrestrial animals, and there aren't any truly terrestrial tetrapods until we get into the immediate neighborhood of the amniotes. According to Romer, in this group -- seymouriamorphs, diadectomorphs, and so on -- the septomaxilla is usually found attached to the lacrimal in the posterior corner of the naris.
 However reasonable this may be, it is not entirely clear that Romer is correct. So, for example, Limnoscelis bears a Type B septomaxilla of classic morphology. It is entirely internal, clearly ventral, has minimal (if any) contact with the lacrimal, and bears a robust dorsal process which would have done justice to Cyrano de Beregerac -- had he been, for example, a temnospondyl.
However reasonable this may be, it is not entirely clear that Romer is correct. So, for example, Limnoscelis bears a Type B septomaxilla of classic morphology. It is entirely internal, clearly ventral, has minimal (if any) contact with the lacrimal, and bears a robust dorsal process which would have done justice to Cyrano de Beregerac -- had he been, for example, a temnospondyl.
CONTINUED ON NEXT PAGE
checked ATW050520