 The Standard Condition
The Standard Condition| Bones: Dermal Bones | ||
| The Vertebrates | Mandibular Series: Dentary |
| s ("Cladograms") |
||||||
| Vertebrates Home | Vertebrate | Vertebrate | Bones | Time |
| Mandibular Series
|
The Standard ConditionTo our right is the jaw of Bob, the standard critter with the Standard Condition. This particular avatar of Bob is so unlike anything that ever lived that we must exercise extra caution. Know ye that Bob is a member of the taxon Didactomorpha, meaning that this jaw is designed to support points of view, rather than points of teeth. However, with that said, know also that in this case, the Standard Condition is extremely standard and applies generally to more or less everything from Cheirolepis on except for a few durophagous, or otherwise pathological forms, and the usual lot of osteological outcastes and untouchables -- the mammals.
The dentary has had minor ups and downs in vertebrate phylospace. The dentary almost always covers the outside of the mandible and holds the outer marginal dentition. Much of its life among tetrapods consists of a slow war of attrition against the coronoids, splenials and prearticular internally and the angular and surangular externally. Generally speaking, the dentary has made gradual and irregular progress in displacing these bones, although it has acheived decisive victory only among the mammals. At the level of Bob, roughly the very basal Amniota, the dentary was carrying all before it on the outside of the jaw. The entire anterior part of the outer jaw was already covered by the dentary. Posteriorly, it was encroaching on the coronoid process and taking large bites from the former territory of the splenial(s) and surangular.
However, these victories were somewhat hollow, or at least the jaw was. The inner surface was still controlled by the archaic splenial, sometimes multiple splenials, as well as the coronoids. The latter still competed with the dentary as the main tooth-bearing elements of the mandible.
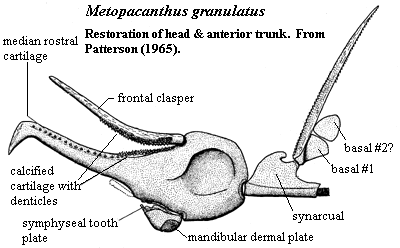 The developmental source of the dentary is quite clear. Like most dermal bones, it began by contour ossification of structural cartilage. Patterson's (1965) restoration of Metopacanthus, a Jurassic myriacanthoid chimaera, provides us with one possible model of how the dentary may have begun. Here, the working parts of the jaw are all cartilagenous, but a small plate of dermal bone has developed at the bottom of the jaw, either for muscle attachment or to guard against mechanical stress and abrasion.
The developmental source of the dentary is quite clear. Like most dermal bones, it began by contour ossification of structural cartilage. Patterson's (1965) restoration of Metopacanthus, a Jurassic myriacanthoid chimaera, provides us with one possible model of how the dentary may have begun. Here, the working parts of the jaw are all cartilagenous, but a small plate of dermal bone has developed at the bottom of the jaw, either for muscle attachment or to guard against mechanical stress and abrasion.
All members of the mandibular series may have began in this fashion (except the articular, which is not a dermal bone). However, the dentary was a bit different because it early assumed the role of supporting the most marginal tooth row. As it turned out, this was an excellent career move, because most major terrestrial vertebrate groups increasingly came to rely on this dentition alone.
However, it may not have happened in this fashion. In the Middle Paleozoic, a range of competing designs existed which might have given rise to the dentary. All are found in the Acanthodii -- another indication of the possible paraphyly of this group. Many acanthodians bore teeth directly on the Meckelian cartilage, which was ossified in most cases (like placoderms), but may have remained cartilagenous in others (like sharks). The "typical" condition, if such a thing exists for acanthodians, seems to have involved a mixture, with the Meckelian ossified both proximally and distally, but remaining cartilaginous in the middle. However, regardless of the ossification state of the Meckelian, most acanthodians also developed dermal bones on the mandible, which might have developed denticles. Any of these might be homologous or at least analogous to the early dentary. However, it is striking that none of these dermal bones are presently known to have supported teeth. Long 1995).
Among the Climatiiformes, the Meckelian is ossified and bears denticles, but the main tooth-bearing function is taken over by an (unnamed?) "dentigerous jaw bone" which lies along the apex of both the upper and lower jaws. Janvier 1996). This is not an ornamented external dermal, but appears to be a special purpose novelty. This, also, is a possible origin for the dentary. However, this model suffers from the opposite problem: none of these bones are known to have developed any significant degree of external exposure. Accordingly, we are forced to retire in confusion, without a clear idea whether the dentary was a normal ordinary dermal bone exapted to hold teeth or a dentigerous bone which aspired to a front office job.
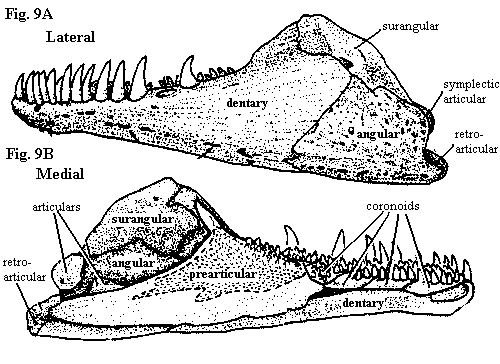 However it began life, the dentary "appears full blown," albeit from the head of Cheirolepis rather than Zeus. Both branches of the Vertebrata at the actinopterygian
- sarcopterygian split inherited the Standard Condition. Among the ray-finned fishes, this was maintained at least through the living neopterygian Amia, which is shown at right. The sarcopterygian fishes developed a tendency to minimize the dentary, but trend was reversed with the evolution of land-dwelling tetrapods.
However it began life, the dentary "appears full blown," albeit from the head of Cheirolepis rather than Zeus. Both branches of the Vertebrata at the actinopterygian
- sarcopterygian split inherited the Standard Condition. Among the ray-finned fishes, this was maintained at least through the living neopterygian Amia, which is shown at right. The sarcopterygian fishes developed a tendency to minimize the dentary, but trend was reversed with the evolution of land-dwelling tetrapods.
Despite the structural advantages of a one-piece jaw, most vertebrates have kept the Standard Condition because it constitutes a workable compromise for any organism with an essentially kinetic jaw mechanism. To explain, Bob, and most other vertebrates apply most of the force of the jaw-closing muscles when the jaws are wide open, relying on momentum to follow it through. This is sound strategy, because it makes efficient use of the adductors, and adductor musculature has probably been the most important limiting factor for tetrapods. Unless one wishes to go to the absurd extravagence of evolving molars, the main thing for a predator is to place nutritionally appropriate organisms (or portions thereof) in the oral cavity and swallow them as quickly as possible. Anything else is essentially waste motion. Thus, the range of action of the adductors should be from mouth open to mouth closed. As muscles work best in the middle of their range of motion, it follows that an efficient adductor system will apply most force to the jaw when the mouth is still fairly wide open.
This is efficient, but not entirely safe. As anyone knows who has ever bitten down hard on something that wasn't there, a kinetic system has certain disadvantages when the cushioning medium of water is absent. Thus, it made sense for land animals to retain a flexible, multi-element jaw with its hollow or cartilagenous core. This design permits a long but lightweight jaw, lots of surface area for teeth, and the flexibility to avoid the worst consequences of miscalculation. Even lineages which have departed from this model have sometimes found it expedient to return to the original inspiration. Review, for example, the jaw of Tyrannosaurus, which differs from Bob's jaw only in its adaptations for great mass. Note particularly how the immediate ancestors of Tyrannosaurus reinvented the multiple coronoids in the guise of a "supradentary".
We might speculate on why the result was different for mammals. Mammals differ because they expect their teeth to do the work of a gizzard or large, muscular gut. Unlike some mammalian adaptations, molars are probably not simply a matter of caprice or willful indifference to the legitimate interests of anatomists. A warm-blooded creature has to eat a lot. Since internal processing takes a long time, one either has to develop a gut of truly Brobdignagian proportions or find a way to process food outside of the stomach. Since McDonald's was unavailable in the Mesozoic, it was necessary for the early mammals to pre-process their own food. Molars were thus helpful co-conditions for the development of a fully warm-blooded lifestyle. Oral food processing, in turn, meant that muscles evolved which worked while the mouth was nearly closed. With less need for a kinetic jaw, this constraint on the growth of the dentary was removed. ATW021223.
checked ATW050520
{kind=link}