|
|
Bones: Dermal Bones |
| The Vertebrates |
Facial Series: Overview |
The Facial Series
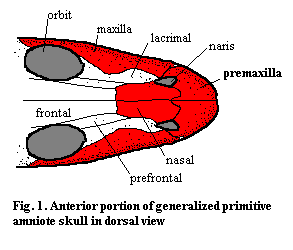 The facial series of the dermatocranium consists of the maxillae, premaxillae, and nasals. Where present, the series also includes the septomaxillae, small bones normally located under the premaxilla which define the internal shape of the external nares. For lack of any better place to put them, the facial series also includes a variety of small rostral bones which occur in various taxa under various different names. From a functional point of view, the facial series defines the rostral region of the skull, some or all of the distal and internal outline of the nares, and the primary tooth-bearing units of the upper jaw. However, this generalization is subject to many exceptions, and should not be taken too literally.
The facial series of the dermatocranium consists of the maxillae, premaxillae, and nasals. Where present, the series also includes the septomaxillae, small bones normally located under the premaxilla which define the internal shape of the external nares. For lack of any better place to put them, the facial series also includes a variety of small rostral bones which occur in various taxa under various different names. From a functional point of view, the facial series defines the rostral region of the skull, some or all of the distal and internal outline of the nares, and the primary tooth-bearing units of the upper jaw. However, this generalization is subject to many exceptions, and should not be taken too literally.
In fact, the facial series does not appear to behave as a strong unit of phylogenetic anatomy. We cannot make generalizations about the evolution of the facial series in the same way as we might speak of evolutionary trends for organ systems or individual bones. The series is simply a convenient way to break up the long list of dermal bones as an aid to memory.
This, in itself may be an important datum. These bones clearly articulate with each other and must be coordinated, at least developmentally, in order to do so. Yet the way in which they fit together does not appear to be under stringent control, or at least under no different control than may be explained by the selective forces acting on the individual bones.
Before running too far with this speculation, it is important to point out some exceptions. The teleost jaw, described elsewhere represents a case in which the development of the premaxilla as the principle tooth-bearing element is part of the more complex structural evolution of the protrusible jaw. Similarly, the concerted retooling of the rostrum towards consolidation in various different fossorial forms, or towards radical kinesis in advanced snakes, appear to represent selection acting on the entire facial series as a unit. However, these exceptions can be explained as cases in which the "normal" (whatever this means) degree of movement between the functional units of the skull has been eliminated in order to deal with the extraordinary stresses imposed by underground, fossorial life. In the case of snakes, and probably mammaliforms, the lineage has rediscovered terrestrial life after a period of mainly fossorial evolution. In these cases, the changes wrought by fossorial specialization have had irreversible effects, with the result that the skull responds in a more unified fashion to selective pressures. Thus, the specialized teleost jaw is the only true exception.
This pattern is consistent with a genetic tale which goes something like this. Once upon a time there were fishes that developed dermal bones as plate armor. Initially, the main selective pressure was to find a pattern of ossification centers which made it convenient, from a developmental perspective, to cover whatever needed to be covered -- normally the braincase, jaws, and sensitive areas around the sensory organs of the head -- with minimum fuss and bother during embryogenesis. In these far-off days before Cheirolepis, this meant that the number and location of dermal ossifications would change almost as frequently as the shape of the head or the type of food preferred. The centers simply grew until they encountered other osteocytes and then quit.
However, anything worth doing consistently is worth doing genetically. By the Late Silurian, a number of different kinds of fish happened on the practice of using dermal bone for secondary functions which required more consistency: bearing teeth being probably the first and best example. This required a greater degree of gene-level control. Dermal bones could no longer develop simply in response to the shape of the underlying epidermis, but began to respond directly to specific genetic signals from neural crest and mesenchymal messengers. Now the best way to ensure consistent and coordinated obedience to such signals is to give the signals early to a single population of stem cells and let them execute the program, with only occasional developmental nudging. Thus, the centers of dermal ossification came to be identified with discrete structural roles. Once even one dermal bone becomes "dependable" in this developmental sense, the bones around it become "dependable" to some degree, since they will stop growing at the defined boundaries of the fixed bone. This means that the surrounding bones are more fixed and consistent, thus more consistently able to take up secondary roles which, in turn, will tend to bring them under genetic control.
If correct, this explains why various different gnathostome lineages developed distinct, but rather different, patterns of dermal bone or abandoned dermal bone altogether by the mid-Devonian. The tooth-bearing function acted as a catalyst for genetic control of the entire dermatocranium. Granted, the sample size is not large: a half dozen different placoderm lineages, the similar, but not identical, actinopterygian and sarcopterygian patterns. But only one non-gnathostome group shows this trait, the possible exception being an advanced group of pteraspidiforms which seem to have developed a fairly consistent, if rather simple, pattern of dermal bones. Even here, the exception may prove the rule. Basal heterostracans have an upper and lower head shield, plus a rather variable set of small plates between them. The excurrent duct for the gills forms generally at the posterior limit of these small plates. While nowhere near as precise a function as tooth-bearing, the formation of this duct might have acted as a nucleus for spreading genetic control of the dermatocranium in a similar, if less stringent way. In fact, the multi-part dermal skeleton remains more or less restricted to a sort of medial belt defined by the gill slit and associated lateral "fins", the upper margin of the mouth, and the rostrum.
Thus the concept of the bone as a unit of evolutionary change has a possible basis in real biology in a way which is less applicable to bigger or smaller anatomical units. As we have seen with the premaxilla, further stabilization may occur when the same bone is subject to multiple genetic controls. However, selection works on larger units only in extreme circumstances which force simultaneous changes in multiple areas of the skull.
Specific Bones:
1. Internasal
2. Premaxilla
Minor Bones:
1. Internasals
A variety of different bones are referred to as "internasals." It is almost certain that these elements are not homologous. As discussed elsewhere, the premaxilla originally functioned largely as a second tooth-bearing bone in the upper jaw. In very primitive gnathostomes, the space between the margin of the jaw and the nares was filled with a number of small rostral bones. In this group, the internasals are generally paired small bones of this type just distal to the nasals. Unfortunately, the term is also used to describe a teleost bone of dermal origin which forms part of the braincase, with a position which seems to be analogous to the mesethmoid in advanced mammals.
In tetrapods, a number of groups occasionally produce small, unpaired bones between the paired nasals which may also articulate with the premaxilla. See, for example, Phylogeny of stegocephalians. Ichthyostega has a central bone of this type. Possibly, these are homologous with the teleost bone of the same name. Such bones are occasionally observed even in humans. Virtual Hospital: Nasal Bone. Thus, the phylogenetically random appearance of internasals in tetrapods may simply be the sampling of random individual variation, or random genetic drift of a minor mutation which has no immediate selective consequences. In fact, the most interesting point may be that there do not appear to be any significant consequences. This is entirely consistent with the general observation that there is at least a bit of slack in the fit of bones before selection becomes significant. ATW 010115
checked ATW050523