|
|
Bones: The Braincase |
| The Vertebrates |
Braincase Overview |
The Braincase: Overview
First Iteration: General Layout & Vague Concepts
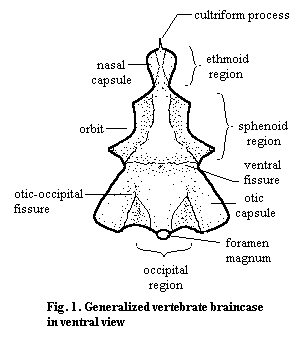
The individual bones of the braincase are primarily embryological units. In the adults of most vertebrates, the braincase is largely fused into a single solid mass. The braincase may be bone, cartilage, or (most commonly) mixed. However, the various embryonic cartilages and centers of ossification are not separated by obvious sutures in the manner of the dermal bones, and there is rarely any marked kinesis between sections of the brain. Nevertheless, there are certain more or less persistent regions and landmarks that are frequently mentioned. These are illustrated schematically in Figure 1.
It should be emphasized that braincases come in a vast array of shapes and sizes. The otico-occipital boundary is sometimes almost seamless. Likewise the sphenethmoid area is frequently a single unit. It is almost always relatively easy to distinguish the otico-occipital and sphenethmoid units, although the ventral fissure is not always present. By way of example, see the braincase of Trilophosaurus from Parks (1969). Fig. 2.
It is interesting, as well as a useful aid to memory, to note that the principal braincase regions are each strongly associated with different sensory functions. The occipital region is defined by the bones of the occiput (the basi-, ex-, and supra-occipitals) which surround the foramen magnum. This is the point where the notochord and the sensory (and motor) nerves from the body enter the brain. The otic capsule partially surrounds the semi-circular canals of the labyrinth, which sense motion and orientation, as well as the organs of hearing in tetrapods. The sphenoid region is essentially defined by the orbit, and the ethmoid region is associated with chemosensation.
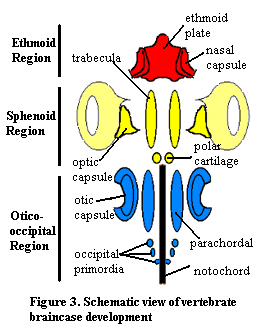 Vertebrate braincase development is relatively stereotyped, even if the final result is extremely variable. Neural crest cells migrate into the head region lateral and anterior to the notochord and condense into a pair of elongate cartilage bars, the trabeculae or prechordal bars, and a pair of small polar cartilages. A similar pair of parachordal cartilages develop posteriorly from mesoderm, flanking the notochord. Occipital cartilages form just posterior to the parachordals. Later in development, the bars widen laterally and extend up around the sides dorsally to form a trough. An ethmoid plate forms anteriorly. The embryonic composition of the ethmoid plate is not known with certainty.
Vertebrate braincase development is relatively stereotyped, even if the final result is extremely variable. Neural crest cells migrate into the head region lateral and anterior to the notochord and condense into a pair of elongate cartilage bars, the trabeculae or prechordal bars, and a pair of small polar cartilages. A similar pair of parachordal cartilages develop posteriorly from mesoderm, flanking the notochord. Occipital cartilages form just posterior to the parachordals. Later in development, the bars widen laterally and extend up around the sides dorsally to form a trough. An ethmoid plate forms anteriorly. The embryonic composition of the ethmoid plate is not known with certainty.
In addition, a pair of sensory capsules develops adjacent to each set of axial cartilages: the nasal capsules, optic capsules and the otic capsules. Later in development, all of these growing regions of cartilage fuse. The product of this fusion is the chondrocranium.
Ancestrally, the chondrocranium was an open-topped pan that housed the brain in a trough chondrichthyans have a chondrocranium that is only slightly modified from this condition. The dorsal surface closes off medially to form a roof over the brain. In bony vertebrates, endochondral bones form from centers of ossification in the chondrocranium, although much of the braincase often remains cartilage. In teleost fish, and occasionally in other groups, separate ossifications occur in the ethmoid region. In mammals, the ethmoid ossifies to form the turbinals, convoluted bones in the nasal cavity that are covered by olfactory sense organs. Parts of the sphenoid region frequently ossify to form the basi-, orbito-, pre-, latero-, etc. sphenoid bones. These, along with the basioccipital, form the platform on which the brain rests. There are several ossification centers in the otic region, notably the prootic and opisthotic. In mammals, this region ossifies into a structure called the petrosal, which houses the inner ear. Finally, around the foramen magnum, these bones develop as the supra-, para-, and basioccipitals. See The Occiput.
 The anatomical relations of the adult braincase are complex and somewhat variable. For simplicity, this discussion is restricted to the more conventional gnathostomes. Dorsally, the braincase and the brain are covered by the dermal bones of the vault series (frontals, parietals, and so on), or the equivalent dermal plates in placoderms. The chondrichthyans, of course, lack dermal skull bones, and here the braincase itself grows up over the brain and forms a completely enclosed capsule. Ventrally, the basioccipital (the ventral member of the occipital series) lies on or over the parasphenoid and pterygoid of the dermal palatal series. Again, sharks are the exception. The shark braincase is perhaps the major structural anchor of the skull and, in primitive forms, it supports the palatoquadrate directly. The occipital series is itself the main support of the posterior skull, and it also serves to bind the braincase to the notochord, primitively the main axial support of the entire organism.
The anatomical relations of the adult braincase are complex and somewhat variable. For simplicity, this discussion is restricted to the more conventional gnathostomes. Dorsally, the braincase and the brain are covered by the dermal bones of the vault series (frontals, parietals, and so on), or the equivalent dermal plates in placoderms. The chondrichthyans, of course, lack dermal skull bones, and here the braincase itself grows up over the brain and forms a completely enclosed capsule. Ventrally, the basioccipital (the ventral member of the occipital series) lies on or over the parasphenoid and pterygoid of the dermal palatal series. Again, sharks are the exception. The shark braincase is perhaps the major structural anchor of the skull and, in primitive forms, it supports the palatoquadrate directly. The occipital series is itself the main support of the posterior skull, and it also serves to bind the braincase to the notochord, primitively the main axial support of the entire organism.
The problem area, in terms of evolutionary engineering, appears to have been lateral support. The dorsolateral portion of the skull is dominated by the jaw adductor muscles. The location, size, orientation and mechanical advantage of these muscles has probably been more critical to vertebrate evolution than any other single body element, the brain included. As humans, our cultural progress has often been driven by speech. But, in evolutionary terms, its how you close your mouth that counts. This creates a serious evolutionary conflict: how can we provide lateral stability for a big, delicate brain sitting in the middle of big, active, important muscles whose size and orientation is critical to survival?
Evolution has devised any number of solutions to the problem, and it would be impossible to cover even the more significant approaches. One of the most frequent designs has involved a posterolaterally directed brace, frequently through the stapes, which extends roughly from the level of the braincase's center of mass down and back towards the heavily stabilized occiput. Another, more radical, design is the peculiar system of the always-aberrant mammals. They circumvent the entire issue by placing the jaw muscles largely outside the dermal bones. This lets the brain get very big indeed without losing stability, but it requires a reoriented, shorter jaw with very little snap. This, in turn, has only been successful because mammals have unique teeth which can process food by grinding, a process driven by the masseter muscles rather than the classical jaw adductors. Whatever the solution, it is important to keep track of how these paramount issues interact: brain size and stability vs. jaw musculature.
Second Iteration: Schematic Guide to the Brain of Bob
By definition, Bob the Basal Amniote has the Standard Condition -- in this case the standard brain. While Bob is not a creature of towering intellectual prowess, we nonetheless value his companionship because of his congenial disposition and straightforward anatomy. Even so, we are perhaps not yet ready to tackle a real braincase head on, as it were. Instead, we will first describe the state of his mind as a schematic, or rather two of schematics of increasing complexity. At this stage, we will restrict ourselves to bones. This, for all practical purposes, eliminates the unossified front half of the braincase.
 The thing at right which looks like a badly drawn dhow is in fact a badly drawn amniote braincase. Only the basic osteology is shown. We also provide an occipital view for reference. Beginning with the supraoccipital and moving roughly clockwise, we'll go through the essential information about each of the main structures.
The thing at right which looks like a badly drawn dhow is in fact a badly drawn amniote braincase. Only the basic osteology is shown. We also provide an occipital view for reference. Beginning with the supraoccipital and moving roughly clockwise, we'll go through the essential information about each of the main structures.
The supraoccipital is essentially an unpaired dermal bone of the occiput. It is a bone found only in tetrapods and develops by ossification of a membrane joining the two otic capsules. The supraoccipital contacts the parietals of the skull table dorsally and the exoccipitals ventrally. It usually forms at least the dorsal edge of the foramen magnum.
The exoccipitals are paired bones of the occiput. They derive from the neural arch elements of embryonic vertebrae which have been incorporated into the braincase. Dorsally, the exoccipitals contact the supraoccipital and the foramen magnum. Ventrally, they contact the basioccipital. The exoccipitals often form part of the occipital condyle.
The opisthotic is the posterior of the two bones making up the otic capsule and is usually fused to the prootic, the anterior otic bone. The opisthotic is an endochondral bone. It is usually the largest contributor to the paroccipital process and to the structures of the middle and inner ear. It normally contacts all of the occipital bones and the prootic, and may contact the basisphenoid. It often forms part of the edge of the foramen ovale, the door to the inner ear.
The paroccipital process may be considered a process of the opisthotic, but the process may be formed by bones in addition to, or even instead of, the opisthotic. The paroccipital process runs horizontally and sometimes dorsally) across the back of the skull. It joins the occiput and braincase to the quadrate, squamosal and the other dermal bones of the "cheek."
The foramen ovale (= fenestra ovalis) is the only one of the various holes in Bob's head which makes our list of basic bits and pieces. It is a membrane covered manhole between the middle ear and inner ear. See, generally, The Ear It is covered by the footplate of the stapes. The foramen ovale is normally located anterodorsal to the paroccipital process, between the prootic and opisthotic.
The stapes is an incarnation of the hyomandibular. Originally, it may have been the main upper element of a gill arch. It later appears as the hyomandibular, an accessory jaw element. In early tetrapods, it becomes a stout bone bracing the braincase against the quadrate. As the paroccipital process took over this function, the stapes was reduced, eventually becoming specialized for hearing as the columella, in sensible amniotes, or the stapes, in mammaliforms. The stapes bears a footplate which fits over the foramen ovale.
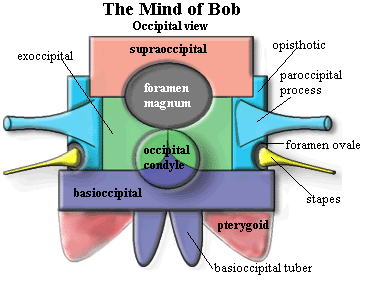 The occipital condyle is a rounded protuberance (or a pair in some taxa) forming the ball of a ball & socket joint by which the head rotates on the neck. It is usually formed by some combination of the basioccipital and the exoccipitals.
The occipital condyle is a rounded protuberance (or a pair in some taxa) forming the ball of a ball & socket joint by which the head rotates on the neck. It is usually formed by some combination of the basioccipital and the exoccipitals.
The basioccipital is an unpaired median bone of the occiput which derives from the centra of embryonic vertebrae which have been incorporated into the braincase. It forms the floor of the braincase under the posterior part of the otic capsule. It contacts the exoccipitals dorsally and almost always forms at least part of the occipital condyle.
The basioccipital tubera are a pair of ventrolaterally directed blobs descending from the basioccipital. They are sometimes simply referred to as "basal tubera." However, the basisphenoid may also bear tubera. Presumably the basioccipital tubera act as attachment sites for ligaments stabilizing the head on the neck.
The basipterygoid processes are (despite the name) processes of the basisphenoid. They act to join the braincase to the palate. In many basal tetrapods and their ancestors, this was a moveable articulation. In most derived tetrapods, it simply staples the braincase to the palatal bones.
The pterygoid is not, of course, a braincase bone. It is the dermal palatal bone which grew up over the old palatoquadrate and eventually took over many of its functions. It is a very complex and interesting bone which, fortunately, we can skip over for present purposes. We have more than enough complex and interesting stuff to go
'round just dealing with the braincase.
The basisphenoid forms the floor of the braincase anterior to the basioccipital. Ventrally it is covered by a dermal bone, the parasphenoid. The fusion between these two is so close that some workers refer to the complex as the "parabasisphenoid." The basisphenoid gives rise to the basipterygoid process and other structures dealt with elsewhere.
The parasphenoid is the dermal bone normally found fused to the basisphenoid on its ventral surface. The parasphenoid generally extends far anteriorly on the midline of the palate as a narrow cultriform process. The upper surface of the parasphenoid (and the vomers) may be associated with the olfactory tracts. However, the anterior braincase is normally unossified in tetrapods such as Bob.
The epipterygoid is another misnomer. Like the quadrates, the epipterygoids are ossified portions of the palatoquadrate (the original upper jaw which, like the hyomandibular, is homologous with an upper gill arch segment). They often appear to arise from the pterygoid, but do not. The epipterygoids are the true, old stuff of the palatoquadrate, while the pterygoid is but common dermal bone with pretensions. In fact, the epipterygoids are the original braincase articulations of the palatoquadrate. They demonstrate this ancient nobility by rising up in a graceful curve to reach the bones of the skull roof, like the last remaining columns in the abandoned temple of some forgotten god. [1]
The prootic is the anterior of the two endochondral bones making up the otic capsule. It is usually fused to the other otic capsule component, the opisthotic. The extent and geometry of the prootic are quite variable. In addition to its fusion with the opisthotic, the prootic may contact the basisphenoid and any of the elements of the occiput. ATW031205.
Third Iteration: Some Landmarks
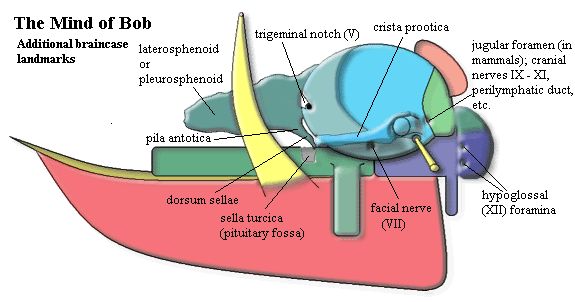 Having disposed of the osteology, we add some landmarks. Here, we are hampered by vague, shifting, and inconsistent nomenclature as well as the more usual obstacles -- among which lack of artistic ability ranks prominently. Nevertheless we have persevered in order to illustrate some critical details, as well as to familiarize the reader with some of the prestigious optional features which are available in luxury models of the tetrapod braincase. Again, we will begin in the upper right hand (posterodorsal) corner and work clockwise.
Having disposed of the osteology, we add some landmarks. Here, we are hampered by vague, shifting, and inconsistent nomenclature as well as the more usual obstacles -- among which lack of artistic ability ranks prominently. Nevertheless we have persevered in order to illustrate some critical details, as well as to familiarize the reader with some of the prestigious optional features which are available in luxury models of the tetrapod braincase. Again, we will begin in the upper right hand (posterodorsal) corner and work clockwise.
The first feature is called by a variety of names, such as jugular foramen, vagus foramen, or perilymphatic foramen, depending on the taxon and anatomical features of interest. Nearly every tetrapod has some sort of major foramen located on the boundary between the exoccipitals and the opisthotic, near the base of the paroccipital process.
The basioccipital may bear several small foramina for the hypoglossal (XIIth) nerve near the base of the occipital condyle.
Anterior, and usually a bit ventral, to the fenestra ovalis is a foramen for the facial (VIIth) nerve. The foramen is often accompanied by a groove, fossa, or what-have-you for the palatine branch of the nerve.
Just anterior to the otic capsule is perhaps the most complex and ancient area of the vertebrate brain. This is the boundary between the embryonic trabeculae and parachordals, the ancient anterior terminus of the notochord, the transition from anterior trabecular prechordal ectoderm to posterior neural crest tissue, the site where palatal (stomodeal) ectoderm invades the brain embryonically, the location of the pituitary, the origin of the optic nerve (II) and critical optic muscles, the seat of the basipterygoid process, the division between forebrain and mesencephalon, and so on. In short, almost everything that distinguishes the vertebrates begins right there, in a suite of features separated only by millimeters. We will mention only a few of these landmarks, but the critical importance of this region cannot possibly be overstated.
The basisphenoid bears a deep, central depression at this point called the sella turcica or pituitary fossa. The sella contains the pituitary (or hypophysis), a structure formed cooperatively of brain tissue and palatal ectoderm which has invaded during development via Rathke's pouch. This is taken up in detail elsewhere. A small sheet of bone rises up vertically at the posterior margin of the sella turcica. This is the dorsum sellae. The right and left corners of the dorsum sellae are usually prolonged anterodorsally. These are the right and left pilae antotica, called the posterior clinoid processes in mammals. The pilae may disappear during development, remain cartilaginous, or ossify in a number of ways. Here, we have shown the pilae ossifying dorsally, as an irregular sheet of bone running roughly between the pila antotica and a similar band of cartilage which comes off the top of the otic capsule, the taenia marginalis. In this position, the bone is referred to as a laterosphenoid or pleurosphenoid In other taxa, a sheet of bone develops ventrally, between the pila and the basisphenoid. In that case, the bone is a presphenoid.
The prootic is quite often emarginated on its anterior border. This is the trigeminal notch which, not surprisingly, contains the exit of the trigeminal (Vth) nerve.
Finally, particularly in lepidosaurs, the prootic bears a lateral crest which runs anteriorly from near the base of the paroccipital process, over the facial foramen, to the anterior margin of the otic capsule. When present, this crest, the crista prootica, is a particularly good landmark since it is quite easy to spot. ATW031206.
Mental Status Examination
This is as far as we can fruitfully go at the moment. We have not discussed the unossified anterior portions of the tetrapod brain, the brains of fishes of all kinds, or the innermost cavities, such as those between the otic capsule and the basicranial bones. Some of these areas are discussed in connection with particular taxa or particular bones. However, this is a reasonable place to stop for the moment, since the information above is all that one really needs to know for most purposes. ATW031206.
[1] Sure, it's an idiotic conceit! But you're not likely to forget it, are you?
"Headnotes"
Dermosphenotic: The dermosphenotic is a dermal bone in the otic region of many actinopterygianfishes, perhaps including groups as early as the guildayichthyiforms. It is typically found at the posterodorsal corner of the orbit and bears a portion of the postorbital sensory canal (sometimes referred to as an extension of the infraorbital canal). The neuromasts of this canal are the only ones to be innervated by the otic branch of the facialis (VIIth) nerve, or by its superficial ophthalmic branch. It typically articulates, or even fuses, with the supraorbitals and may appear to be serially homologous with them. The dermosphenotic may also articulate with the postorbital or infraorbital series and with the nasals and/or frontals. Posteriorly, the dermosphenotic articulates with the dermopterotic, to which it is developmentally related. The dermosphenotic overlies the autosphenotic, a bone of the neurocranium; and the two are frequently fused to form the sphenotic. See Adriaens & Verraes (1998); Lund 2000).
checked ATW060127