
| Chelonii | ||
| The Vertebrates | Casichelydia |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Anapsida ?
│
└─○Chelonii
├─Odontochelys
└─┬─Proganochelys
├─Chinlechelys
└─○Casichelydia
├─Palaeochersis
├─Australochelys
└─┬─Proterochersis
└─○Cryptodira
├─Kayentachelys
└─┬─Meiolanoidea
└─┬─Kallokibotion
└─┬─Pleurodira
├─Paracryptodira
└─Eucryptodira
|
Contents
Overview |
Taxa on This Page
|
|
| The ancestral cryptodire Kayentochelys aprix, Early Jurassic of North-west Pangea, life reconstruction by Nobu Tamura, Creative Commons Attribution from via Wikipedia |
In the eighteenth century, all Chelonian species were included in Linnaeus' single genus, Testudo. By the early 19th century, this genus had been subdivided into further genera. Higher category classifications were at first based on habitat-related features; but as morphologic information became available new higher taxa such as the suborders pleurodires (side-neck) turtles and the cryptodires ("hidden"-neck) turtles (both named by Cope in the 1860s), were recognized. (Gaffney, 1984 )
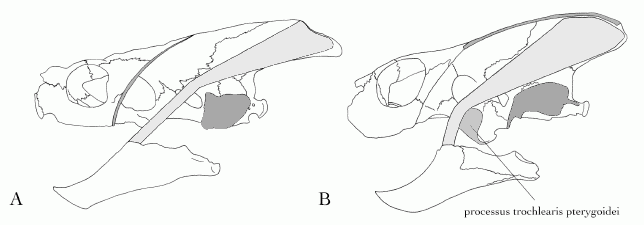 |
| Jaw closure mechanism, traditionally used as one of the ways of classifying chelonians. A, the cryptodire Chelydra serpentina , showing the main adductor musculature (light grey) is redirected by the otic chamber (dark grey). B, the pleurodire Elseya dentata, in which the jaw muscles are redirected by the processus trochlearis pterygoidei (medium grey). Diagram modified from Anquetin, 2009 p.32, in turn modified from Joyce 2007 p.73 |
These two groups differ in a number of ways, including the manner in which the pelvis is attached to the shell, the method used for retracting the neck, and the use of different "pulley" mechanisms to give additional mechanical advantage to the muscles closing the jaws.
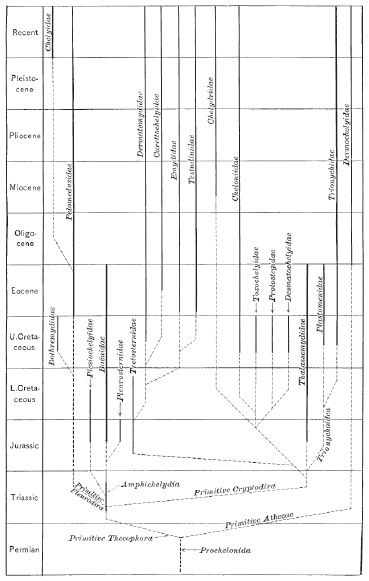 |
| Earliest phylogeny of higher taxa of living and extinct turtles, published by Hay (1908). From Iverson et al 2007 p.86. The hypothetical Permian branching of ancestral testudines (here, "Prochelonida") into Thecophora and Athecae reflects the popular at the time but now long rejected view that the Dermochelyidae or leatherback sea turtles are distinct from all other chelonii because of the absense of a bony shell. It is now known that both Dermochelyids and Cheloniids (hard shelled sea turtles) evolved from a common hard shelled ancestor sometine during the Late Cretaceous, |
At the time, many Jurassic turtles were being rather arbitrarily placed in either the Cryptodira or the Pleurodira. ln 1889 Lydekker erected a suborder Amphichelydia (Paleobiology Database), further elaborated by Baur 1891 and Hay 1905. At this time, there was the tendency to adopt something along the lines of the following classification (modified from Hay 1908 p.25) (Ironically, the distinct position of the Trionychoidea (also in Zittel, Eastman, et al. 1902) would be confirmed a century later by molecular sequencing)
As described by these various authors, the Amphichelydia represented those testudines which had not yet developed the distinctive features of either of the two living suborders, and a taxonomical solution to the problem of many forms that were too far removed from living groups to be identified in one or the other. They were defined for example by a supposed intermediate state between the cryptodire free pelvis and pleurodire condition fused pelvis (Gaffney argues that in these and other regards the Baenidae are actually much closer to the Cryptodiran nature (Gaffney, 1975 pp.407-414)) as well as the skull showing characters that would be expected in the ancestors of Cryptodira and Pleurodira, and a non-retractable neck. Simpson 1938 pp. 251-252 and Romer 1956, pp. 497-498) considered the Amphichelydia as essentially a "horizontal" group (or grade) of intermediate mostly Mesozoic, forms, such as the Pleurosternidae and Baenidae, and including the common ancestors of the two modern groups, the Cryptodira and Pleurodira (Gaffney, 1975 pp.402-6)
An additional suborder, Proganochelydia, was erected by Romer (taxon list) in 1966 for specifically Triassic ancestral forms (Gaffney, 1975, p.407). Eunotosauria sometimes turned up as well, as it was thought at the time that the parareptile Eunotosaurus might be a possible candidate as testudine ancestor (Watson, 1914, Haughton & Brink. 1954 cited in The Paleobiology Database)
This gave the following situation by the 1960s to mid 70s (from Colbert 1969)
A revolution in chelonian systematics occured with Eugene Gaffney's ground-breaking application of cladistic methodology. The aim was to find a testable methodology for determining evolutionary relationships (i.e., phylogeny), along the requirements suggested by philosopher of science Karl Popper, according to whom science should be understood as series of refutable but not confirmable hypotheses. Here Willi Hennig's phylogenetic systematics was selected as the most fruitful approach (Gaffney, 1975 pp.391-2). Hennig had developed cladistics to understand insect evolution, insects have a notoriously poor fossil record. This was one of the first instances of cladistics (phylogenetics) applied to a tetrapod taxon, well represented by fossils. As a result of detailed analysis, Gaffney rejected paraphyletic taxa like Proganochelydia and Amphichelydia, although Proganochelys itself was recognised as an ancestral form. The genera previously placed in the Amphichelydia were relocated among the monophyletic Pleurodira and Cryptodira (however Proganochelydia was retained in Linnean classifications (e.g. Wikispecies). This gave the established consensus from the mid 70s to the mid noughties (Gaffney, 1975, Carroll, 1988, Meylan 2001), as shown by the cladogram on the left
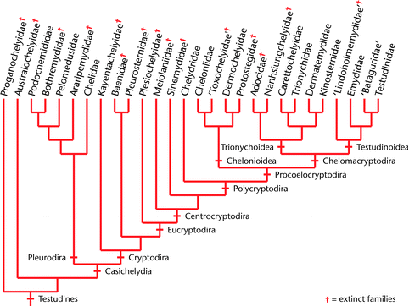 |
| Testudine cladogram, based on Gaffney, 1975 and later papers, showing both extinct and living forms. From the Encyclopedia of Life Sciences, via biogeography of leatherback turtles. |
At the base was Proganochelys . From this ancestral stem (to which were later added additional very basal forms like Australochelys and Palaeochersis), as early as the Late Triassic to Earliest Jurassic, chelonians had already diversified into the two established clades, the Cryptodira and Pleurodira. Neither group had any mechanism for withdrawing the neck under the shell in these basal forms. Both groups independently evolved this defense mechanism much later, during the Cretaceous.
The relation between the Pleurodira, Paracryptodira, and Eucryptidira has become somewhat more complicated following molecular phylogenies. In both earlier and more recent cladistic morphological-based accounts, the Pleurodira are always distinct from the Cryptodira, the two being monophyletic taxa . This is the case regardless of whether there is an earlier Gaffney, 1975, Gaffney and Meylan, 1988) or later (Joyce 2007) divergence between the two groups. Moreover, the Trionychoidea or soft-shelled turtles are nested deep in the Cryptodiran crown group. This has been challenged by certain molecular studies (Krenz et al 2005, Barley et al. 2010) which not only are unanimous regarding the polyphyletic (artifical) status of the Trionychoidea, but also place one group, the Trionychia (= Trionychoidae), at the base of the Cryptodiran family tree (Shaffer, 2009, Barley et al. 2010, p.1190). A few results even raise the possibility that the pleurodires are a specialised branch of cryptodires, and the Trionychia being the most basal group. This would mean the cryptodire condition being the ancestral morphological states and the pleurodire the derived one. It also means that the cryptodire neck retraction mechanism evolved twice. Several cladograms also nest the pleurodire within the cryptodires (and not just within the cryptodires but the eucryptodires) (Joyce 2007 p.56 fig.15, Anquetin, 2009 pp.187f), and this counter-intuitive result may be simply due to convergent characteristics (Joyce 2007 p.53), although the general parallel with some of the molecular trees is intriguing. However whether the cryptodires are paraphyletic or monophyletic vary according to the type of analysis, as shown in the phylograms below. In any case it would seem that these various groups probably emerged through a rapid evolutionary radiation (Danilov & Parham, 2006, p.579; Krenz et al 2005, p.189, but see Shaffer, 2009 for the opposing view) the possibility that the pleurodires are a specialised branch of cryptodires, p.189; ), which is why resolation of the branching order is difficult. Whatever the truth of the situation, chelonian evolution is clearly a lot stranger and more complex then was previously supposed. MAK100902
A few cladograms...
The following diagrams combine two sets of molecular analyses with the Trionychia + Durocryptodira of Danilov & Parham, 2006 and 2008:
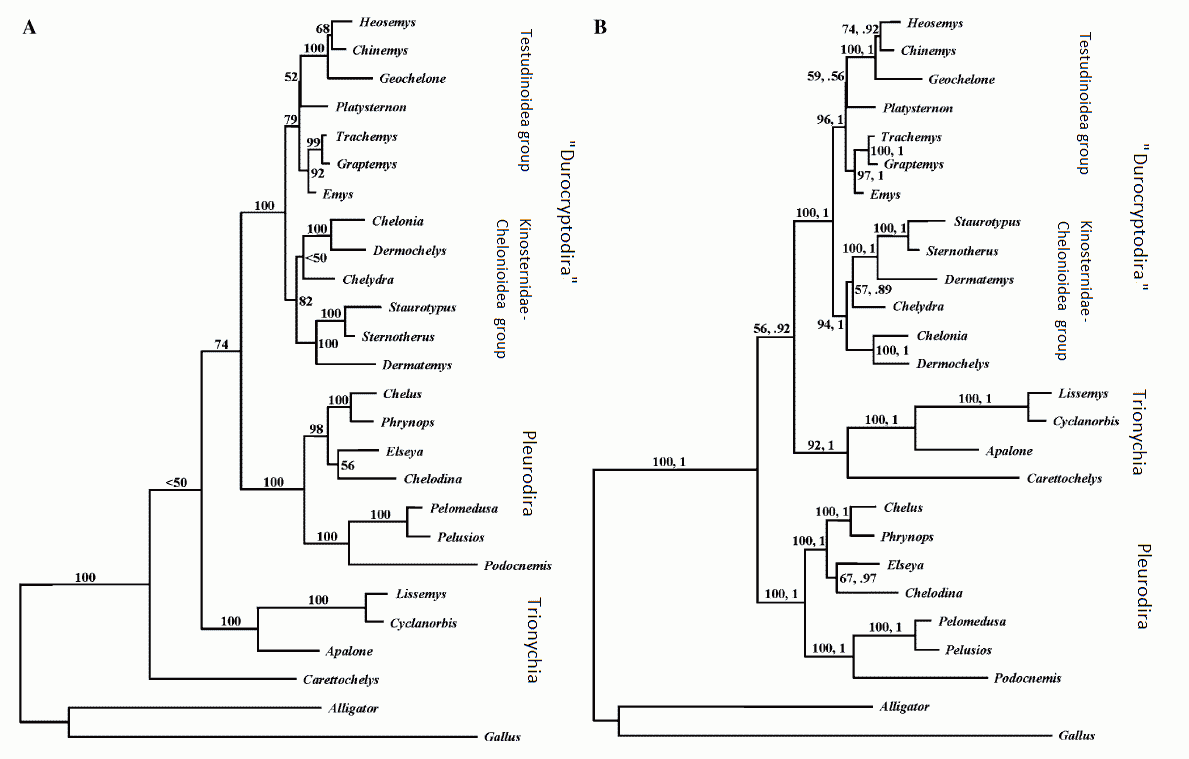 |
| Molecular phylogeny according to Krenz et al 2005, (A) Maximum parsimony phylogram for 2793 nucleotides of RAG-1 ( (Recombination activating gene,a slowly evolving gene which has a very homogeneous base composition across the included taxa (Krenz et al 2005, p.186.)) for 24 turtle species. (B) Maximum likelihood and Bayesian analysis phylogram of RAG-1. Similar results are had using a combined analysis of RAG-1, cytochrome b, and 12S rDNA nucleotides (ibid, p.187 fig.5). |
 |
| Molecular phylogeny according to Barley et al. 2010. (a) Phylogram generated from concatenated phylogenetic analysis of 14 nuclear genes.. Concatenation uses phylogenetic reconstruction is done after the gene sequences are concatenated head-to-tail to form a super-gene alignment, in contrast to the consensus approach where phylogenies are inferred separately for each gene and the resulting gene trees are used to generate a consensus phylogeny (Gadagkar et al 2005, pp.64-5). (b) Species tree generated from BEST phylogenetic analysis of 14 nuclear genes. BEST uses the joint posterior distribution of gene trees in order to estimate a species tree in a multiple locus analysis (Liu & Pearl, 2007) |
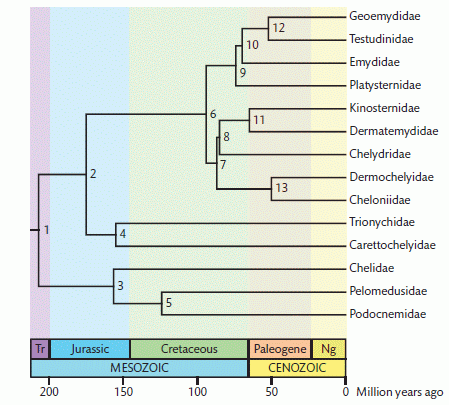 Molecular phylogeny matched with molecular clock (note that molecular clocks aren't the same as molecular phylogeny, although the two are often matched), which supports an early diversion of Pleurodires and Cryptodires (Gaffney, 1975), but is incompatable with more recent analyses and discoveries (Joyce 2007 ,Sterli, 2008, Anquetin et al. 2009), from Shaffer, 2009 (url) and more recent molecular dating (Lourenço et al 2012 p.502). I tend to be sceptical of too much reliance placed on the molecular clock, because it assumes that evolution proceeds at a steady rate. In fact there many obstacles to accurate molecular dating, including not only rate heterogeneity but also use of inappropriate fossils as a source for calibration dates and the fact that fossils will almost always underestimate lineage originations (Parham & Irmis, 2008, p.132) However, the antiquity of the Trionychia (Danilov & Parham, 2006 is supported here. By this dating the Paracryptodires and Eucryptodires would be located between node 2 and node 6
Molecular phylogeny matched with molecular clock (note that molecular clocks aren't the same as molecular phylogeny, although the two are often matched), which supports an early diversion of Pleurodires and Cryptodires (Gaffney, 1975), but is incompatable with more recent analyses and discoveries (Joyce 2007 ,Sterli, 2008, Anquetin et al. 2009), from Shaffer, 2009 (url) and more recent molecular dating (Lourenço et al 2012 p.502). I tend to be sceptical of too much reliance placed on the molecular clock, because it assumes that evolution proceeds at a steady rate. In fact there many obstacles to accurate molecular dating, including not only rate heterogeneity but also use of inappropriate fossils as a source for calibration dates and the fact that fossils will almost always underestimate lineage originations (Parham & Irmis, 2008, p.132) However, the antiquity of the Trionychia (Danilov & Parham, 2006 is supported here. By this dating the Paracryptodires and Eucryptodires would be located between node 2 and node 6
Over the last several years however, this picture has been supplemented and challenged by new cladistic studies. The main thesis here is Joyce 2007 , which supports the earlier but then forgotten work of Dryden 1988. This has been supplemented by new discoveries (Anquetin, 2009, Anquetin et al. 2009, Sterli, 2008), all of which place the pleurodire-cryptodire divide much later, so that a number of basal genera previously considered pleurodires or cryptodires are now understood to be part of a diverse assemblage of transitional stem forms. These intermediate types were placed at the stem of the Chelonian tree, albeit higher up than Proganochelys and company, but predating the Pleurodire-Cryptodire split. Proterochersis and Kayentachelys (previously considered the earliest pleurodire and cryptodire respectively) , along with a host of Jurassic and later forms, now represent a paraphyletic assemblage of basal types. In this revised phylogeny, true pleurodires and cryptodires did not appear until the mid to late Jurassic.
Concerning this, a few more cladograms (or more technically, dendrograms), emphasising stem testudine taxa (refs Mikko's Phylogeny Archive, Joyce 2007 p.4, and Anquetin 2009 p.13), can be offered here:
One reason for the very different phylogenies aquired through Gaffney and his coworkers on the one hand, and more recent authors, is the former consider choosing fewer, well-analysed characters is better than large numbers of poorly analysed characters ( Gaffney et al 1991. The ability of more and more powerful computers to process greater amounts of quantitative data does not necessarily give more accurate trees. Another problem is homoplasy, the independent aquiring of the same characteristics by different lineages. Hence the contradiction between what might be called the Gaffney paradigm and the Joyce paradigm, and the even bigger problem of the disjunction between morphology-based and molecular analyses. MAK110922 130311
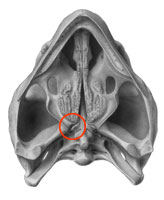  |
| Palate view of skull of Proganochelys (left) and Australochelys (right) showing basicranial articulation. Proganochelys has an open articulation, indicating its primitive status. More advanced chelonians, such as Australochelys, Palaeochersis, and Proterochersis, and all more derived forms, have a fused articulation. Graphics from the Royal Tyrrell Museum - Phylogeny of Turtles - the reasearch interests of Eugene Gaffney |
from the Late Trassic. Cosmopolitan
Testudines : Proganochelys + Chinlechelys + * : Australochelys + Palaeochersis + (Proterochersis + Cryptodira)
Characters: closed interpterygoid vacuity, middle ear with lateral wall present (Gaffney, 1975 p.430), lacrimal formane absent, vomer single,, fusion of braincase and palate (Benton, 2004 p.233)
Comments: "Casichelydia" was coined by Gaffney, 1975 for Pleurodira + Cryptodira. The discovery of a number of new primitive Middle Jurassic species, and the current statistical-computational approach in cladistic methodology, means that a number of taxa that have since been relocated outside the turtle crown group (see e.g. Joyce 2007, Anquetin et al. 2009, and Sterli, 2008 and comment here)
[2] To explain the jargon listed under "characters" above (most of the following is taken pretty much verbatum from Meylan 2001's very readable explanation): Much of early testudine evolutions involved a reduction in the number of bones that make up the skull. The lacrimal bone and lacrimal duct (which is the basic or primitive condition for amniotes) occurs in Proganochelys but not any other chelonians. Likewise, the presence of a pair of vomers seems to be a primitive amniote condition. Again, Proganochelys has two, all other chelonians one. A supratemporal bone is found in primitive tetrapods including Proganochelys, but not in higher testudines. As well as the reduction in the number of bones (which no doubt helps to strengthen the skull), there is also a closing up of the skull which again results in a more solid structure. The moveable articulation of fusion of braincase and palate (or neurocranium and palatoquadrate to use osteological jargonspeak), which is found in generalized amniotes and in Proganochelys is absent in Australochelys and other chelonians (see diagram), where these elements are tightly sutured. Generalized amniotes, such as captorhinids, and also Proganochelys, have a middle ear region that is open at least at the sides (laterally) and the bottom (ventrally). Technically speaking, Casichelydia should include the LCA of Proterochersis and Kayentachelys and all their descendents, which means that it does not include Australochelys. However I have followed the account given by Meylan 2001. Benton, 2004 p.233 gives a cladogram that distinguishes between the node leading to Australochelys and the next one up which is the Casichelydia. But the characteristics he give for each are both covered by Meylan 2001. MAK100917 130120
References: Gaffney, 1975 pp.425, 430; Meylan 2001; Benton, 2004 p.233 .
Links: Palate view of Proganochelys and Australochelys
Early Jurassic of South Africa.; related form (Palaeochersis) from Late Triassic of Sth Am.
Casichelydia : Palaeochersis + (Proterochersis + Cryptodira) + *
Characters: fusion of braincase and palate (see graphic, below left), partial enclosure of middle ear. Retain some palatial teeth and long tail. May or may nor form a monophyletic clade (family Australochelydidae) with Palaeochersis MAK100917 130120
References: Gaffney and Kitching 1994, Gaffney and Kitching 1995, Benton 2000 p.235

Palaeochersis : Palaeochersis talampayensis
Range: Latest Triassic of South America. (Upper Los Colorados Formation, Argentina)
Casichelydia : Australochelys + (Proterochersis + Cryptodira) + *
Comments: Study of forelimb proportions shows that, like the related but more primitive Proganochelys, this was a terrestrial form, indicating a terrestrial origin for early testudines (Joyce & Gauthier, 2004). Included with Australochelys in the Australochelydidae MAK100917
Graphic: Paleoargentina
Size: Length: 70 cm
References: Rougier et al. 1995
Links: Palaeocritti - Palaeochersis talampayensis

Late Triassic of Europe.
Casichelydia : Australochelys + Palaeochersis + (Cryptodira + *)
Comments: many standard pleurodire features, e.g., pelvis fused to shell. For this reason was for a long time considered the oldest known pleurodire (e.g. Gaffney, 1975, Carroll, 1988, Gaffney & Meylan 1988, Hirayama, Brinkman, & Danilov, 2000, Meylan 2001, etc). The current consensus however is that this represents an earlier stem-testudine (Joyce 2007, Anquetin 2009, Sterli, 2010, Pérez-García & Murelaga 2012, etc etc) that predates the Pleurodires. As parallelism seems to be frequent in testudine evolution, it would not be surprising if pleurodire features arose several times independently. MAK130120
Studium Integrale Journal 6/2 (Oktober 1999): Streiflichter (German); Untitled Document; Pleurodira.
010412.
Graphic: Plastron of Proterochersis robusta, photograph by Ghedoghedo, Wikipedia
 Cryptodira : snapping turtles, soft-shells, tortoises & sea turtles.
Cryptodira : snapping turtles, soft-shells, tortoises & sea turtles.
from the Early Jurassic
Casichelyidia : Australochelys + Palaeochersis + (Proterochersis + * : Kayentachelys + Condorchelys + Eileanchelys + (Meiolanoidea + (Kallokibotion + Hispaniachelys + Testudines crown group)))
Head (refer to BIODIDAC): pentagonal in dorsal view, with 2 large temporal emarginations dorsally; parietal covers the crown of the skull, which is markedly domed and tapers to a thin ridge between emarginations; this post parietal process overlies a long post supraoccipital process; occipital condyle below, and overhung by, supraoccipital process; ventrally, the skull has 2 large openings roughly below the emarginations, with a strong pterygoid bar passing between them; post margin of skull formed dorsally by squamosal + opisthotic & ventrally by quadrate + basisphenoid; small medial frontals ant to parietals; broad postorbitals form sides; prefrontal at apex pf rostrum; prefrontal and vomer in contact; (refer to BIODIDAC) upper jaw formed by maxilla w. small premaxilla at tip, both covered in life with keratinous beak; jugal and quadratojugal line up posterior to maxilla; $ jaw adductor tendon passes over process of otic capsule formed by prootic and quadrate; adductor may pass over parietal as well in some species.; $ vertical flange on external process of pterygoid; lower jaw (see BIODIDAC) formed largely by dentary and surangular; dentary has splenial and (post to splenial) a coronoid; retracted neck bent vertically; neck bend by way of specialized condyles – details vary among species. & may have evolved independently; no J forms have ability to retract neck; 2 relatively distinct sacral vertebrae; pelvic girdle sutured (not fused) to both plastron and carapace (see BIODIDAC and shellgirdle). Dominant crown group except in Aus; only Recent turtles in Northern hemisphere.
ATW030531
Comments. Traditionally considered the sister group to Pleurodira (Gaffney, 1975, Joyce 2007) although some morphological findings nest the Pleurodires within the Cryptodires, making the latter paraphyletic group (Danilov & Parham, 2006, and Anquetin, 2009), some molecular research suggests likewise (Krenz et al 2005, Barley et al. 2010). The current nomenclatural tendency now is limit the clade to Crown Cryptodira (= Polycryptodira Gaffney), although not everyone agrees with this form of taxonomy (see notes on "tetrapoda"). To retain consenus with both Evolutionary Linnaean (e.g. Carroll 1988) and Hennigian (via Gaffney) formalism we are using a morphology- (in cladistics-speak "apomorphy") based definition here instead MAK100917 130119
Image: Geochelone carbonaria from Merkblätter.
Links: Turtle, Tuatara, Crocodile Checklist--3; Halsbergers Cryptodira); \Snapping Turtle Page\ - information and photo gallery page; WIEM: Zólwie skrytoszyjne (Polish); Higher Relationships of Testudines; Mongoleus; Order Testudines (Czech & English); Higher Reptile Taxa; Testudines; Cryptodira; Basal Cryptodira Mikko's phylogeny); CRYPTODIRA very nice summary of paleodiversity); APUS.RU | Подотряд Скрытошейные ·черепахи - Cryptodira; Halsbergerschildkröten (Cryptodira)- Kriechtiere a remarkable set of links); Phylogeny of Turtles/Dr. E. S. Gaffney; tierdach.de- Halsberger-Schildkröten German: possibly Best on the Web); Dr. Robert Wochesländer abstracts); home2; Ocean Ambassadors -- Turtle Biology sea turtles, natural history); Testudines Turtle Skull Reproductions; Jomonjin_KameDataBase. ATW030531. Wikipedia
 |
| Kayentochelys aprix carapace, from Anquetin, 2009 |
Range: Early Jurassic of North America (Arizona).
Cryptodira : Condorchelys + Eileanchelys + (Meiolanoidea + (Kallokibotion + Hispaniachelys + Testudines crown group))) + *
Comments: The oldest known cryptodire, based on its trochlear system, other characters, such as palatal teeth, are plesiomorphic with respect to all extant pleurodires and cryptodires (Dryden 1988, cited in Gauthier et al, 1989). As with Proterochersis, the current consensus, based on statistical cladistics, is that it predates the Pleurodire-Extant Cryptodire split. This means that cryptodire features are primitive for stem cryptodires. MAK130119
Links: Kayentachelys.
checked ATW030416, page MAK100917. Page last modified MAK1301020.
All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners
{kind=link}
{kind=link}
{kind=link}