The Rhaetian Age - 1
The Rhaetian Age of the Late Triassic Epoch: 204 to 200 million years ago
Rhaetian - Overview
The Rhaetian is the last of three divisions in the Upper or Late Triassic. It was also the first Triassic Stage to be established (Gümbel 1859, cited in Kozur 2003). The Rhaetian Age succeeds the Norian Age and is followed by the Hettangian (earliest Jurassic). The name is derived from the Rhaetian Alps of Italy, Switzerland, and Austria, where rocks of this age are known. The stratotype (the defining section of rock formation) is the Kössen beds at Kendelbachgraben, Sankt Wolfgang, Austria, where the bivalve Rhaetavicula contorta occurs. [Encyclopædia Britannica]
On land, the Rhaetian saw the continuation of the rule of the dinosaurs among the megaherbivores that began in the late Norian. Other late Norian (Sevatian) floras and faunas seem to have continued with little change; so much so that the Sevatian Stage is sometimes taken to include the Rhaetian as well as the Late Norian [see e.g. Benton 1994]
The most significant thing in the history of life during the Rhaetian was the replacement of typically Triassic marine reptiles (nothosaurs and shastosaurs) with the classic dolphin-like ichthyosaurs and "Nessie"-style plesiosaurs . This change is so dramatic that with the exception of a single genus of placodont ("reptilian walrus"), all Rhaetian marine reptiles known are of the classic "Liassic" (early Jurassic) type.
The Rhaetian age and Triassic Period ended with a mass-extinction (possibly the result of an extraterrestrial impact) that killed off various taxa including some types of plankton, most reef-building organisms, the last of the pelagic conodonts and (assuming they hadn't died out earlier) the straight-shelled nautiloids, all the ceratites, the last of the placodonts, many types of bivalves, a number of amphibian and reptile types, and all non-dinosaurian and non-crocodylomorph archosaurs.
Stratigraphy and Index Fossils
The bivalve Rhaetavicula contorta, which was used to establish the Rhaetian (Kössen Beds) is widespread in extra-Tethyan basins, such as the Germanic Basin, and, therefore, the Rhaetian was also widely used outside the Tethys. However the exact ammonoid definition was for a long time not resolved. This caused problems with the Norian-Rhaetian boundary within the Tethys and in other pelagic sequences, but the Rhaetian Stage was confirmed finally by the Triassic Subcommision in 1993. [Kozur 2003]
Stratigraphically, the base of the Rhaetian is defined as the "near lowest occurrence of ammonite Cochlocera, conodonts Misikella spp. and Epigondolella mosheri, and radiolarian Proparvicingula moniliformis." The end of the Rhaetian (and of the Triassic, and start of the Jurassic) is defined by the "lowest occurrence of smooth Psiloceras planorbis ammonite group." [Gradstein, Ogg, Smith, A Geologic Time Scale 2004 - GeoWhen database - Rhaetian ICS Stage]
Rhaetian Index Fossils - Terrestrial
The Apachean LVF
Dating Rhaetian terrestrial faunas is difficult. Dr Spencer Lucas (ref) divides Triassic tetrapod evolutionary communities into eight land-vertebrate faunachrons (LVF). The last of these, the Apachean corresponds to at the Rhaetian, but may also include the later part of the Norian. It begins with the FAD of the phytosaur Redondasaurus and ends with the FAD of the crocodylomorph Protosuchus. The Redondo Member, which features the type Apachean fauna, is considered tentatively Rhaetian by Lucas 1998, but is referred to as middle to late Norian by both Benton 1994 and Hargrave 1999 This shows the difficulty of providing precise dating. Also, the endemic nature of the Redondo fauna makes it difficult to correlate with the fairly well-dated Stubensandstein and Rhaetian of Europe. For this reason it may not be impossible that the Apachean begins at around the later Middle Norian.
As well as by the Redonda Formation in east-central New Mexico, and the Ghost Ranch Whitaker quarry in the Rock Point Formation of the Chinle Group at Ghost Ranch, New Mexico (however this latter dated magnetostratigraphically as Late Carnian / Early Norian age by Hargrave 1999), the Apachean also includes the upper part of the Cliftonian assemblages, of the Newark Supergroup, the Coloradan Land Vertebrate Fauna of Argentina, and probably some of the fissure fill assemblages in the uppermost Mercia Mudstone Group and lowermost Penarth Group of England, Saint-Nicolas-de-Port and other vertebrate sites in France [Lucas 1998].
Fish
Study of the French Lons-le-Saunier site indicates a change in the marine faunas during the Rhaetian transgression, preferentially affecting the neoselachian sharks, which increase in abundance, and the durophagous bony fishes, which become dominated by the species Sargodon tomicus. [Cuny et al 2000]
Late Norian and Rhaetian Terrestrial Biotas - Tetrapods
As mentioned on the Norian page, the Norian terrestrial fauna represented two distinct and well-defined groups. The Early to Middle Norian shows similarities with the Carnian biota, as well as unique characteristics of their own, but are stratigraphically easily separated from both earlier and later communities. In contrast both large and small (i.e. European "fissure fillings") Late Norian tetrapods are difficult to separate from those of the Rhaetian, and terrestrial stratigraphy is often ambiguous (especially of Gondwana and Asia, where there are no common species with those of the well dated European series). These are usually just noted as "Upper/late Norian - Rhaetian". Dating of fissure fillings is even more difficult.
The main problem here is that unlike the late Carnian through to Middle Norian, the Late Norian and Rhaetian was a period of great provinciality, as far as tetrapods go, and it is rare to find even a single cosmopolitan genus. However, with the exception of North-West Pangean ecological communities, which - assuming the Redonda and Ghost Ranch fauna are of this period - still have a primitive "thecodont" (phytosaur-aetosaur) dominated aspect, all other major terrestrial ecosystems are sauropodomorph dominated. The Late Norian Plateosaurus-Proganochelys Community Type of the Oberer Stubensandstein and Knollenmergel is characterized by the large and abundant herbivore Plateosaurus, a dinosaur that persists into the Rhaetian, as do the plagiosaurs and phytosaurs that co-exist with it. The Plateosaurus community is ecologically very similar both to the Late Norian South African Lower Elliot Formation (protosauropods Melanorosaurus and Euskelosaurus, and early sauropod Antetonitrus), and the Rhaetian Argentinean Los Colorados (diverse large sauropodomorphs). Basal sauropods are also found in the Rhaetian of England (The 9 meter long Camelotia borealis) and the Rhaetian Nam Phong Formation, Thailand (the 6 meter long Isanosaurus). Even the isolated "fissure fillings" were characterized by sauropodomorphs - the small Thecodontosaurus is a very common element.
Ironically, while the evidence of skeletons indicates a continuing dominance by Sauropodomorphs, and the Los Colorados local fauna provides as diverse an assemblage as anything the Norian produced, the footprints of this period tell a different story. If this is anything to go by than the facts are clear - whereas the Norian witnessed the dramatic rise of the Sauropodomorpha, the Rhaetian saw the theropods emerge to dominance; although not yet very large theropods. Rhaetian footprint assemblages are predominantly theropod, and these replace the more diverse Norian assemblages. At this time "all faunas from footprint-bearing horizons are both less abundant and less diverse, and mainly consist of tridactyl bipeds (grallatorids)...Apatopus, the presumed phytosaur trackway, persists from the earlier (Carnian-Norian) section." [Haubold 1986 p.194] . This faunal displacement is especially apparent "in the Chinle Formation and the overlying Glen Canyon Group, in which three toed Grallator type tracks, attributed to early dinosaurs, become increasingly abundant, and increasingly large, as a function of stratigraphic height." [John Stear - Triassic Strata of the Colorado Plateau ]. And according Lockley and Hunt (cited on that webpage)
"Toward the end of the Triassic, in youngest Chinle Group sediments, small-sized Grallator tracks appear in association with a diverse assemblage of other archosaur tracks. Then in some of the very youngest Chinle layers (for example, the Sheep Pen Sandstone) these Grallator tracks become the dominant track type to the virtual exclusion of all other footprint types. We get the impression that the small dinosaurs were taking over. This impression if confirmed by the track record in the overlying strata of the Wingate Formation, where we find more medium-sized Grallator tracks, and very few other track types"
(Lockley and Hunt 1996 p. 103).
Rhaetian terrestrial faunas are with a few exceptions a lot more poorly known than those of the Norian - the following therefore is even more speculative a list than the Norian one.
 |
A representative Sauropodomorph fauna - Late? Rhaetian of South-West Gondwana (La Esuina Formation, Argentina) - showing approximate abundance. Herbivores in outline, carnivores in black
Key: A, Prosauropoda; B, Stagonolepidae; C. Tritylodontidae; D. Protosuchidae; E. Rauisuchids; F. Trithelodontidae; G. Sphenosuchidae; H. + I : Ornithosuchidae; J. Theropoda indeterminate (= ?)
|
Sauropodomorph (Riojasaurus)-Aetosaur Community. By far the best known Rhaetian assemblage is the Los Colorados Local Fauna found in the La Esuina Formation, Argentina. here we see a rich and diverse Sauropodomorph-dominated fauna featuring dinosaurs, other archosaurs, amphibians, turtles, and cynodonts, very much like the Middle and Late Norian assemblages of Europe, Greenland, Thailand, India (all The Prosauropod-Proganochelyid-Aetosaur Community - central-north to mid southern Pangea), and Southern Africa (here referred to as the Melanorosaur-Sauropod (Euskelosaurus) Community - South Pangea). The dinosaurian fauna combines both the prosauropod elements common in the north with the more advanced basal true sauropods of the south. However, the most common animal (Riojosaurus), is a Prosauropod like the northern Plateosaurus. The lack of any common faunal elements with the geographically very close Southern African Lower Elliot fauna indicates either a lot of geographic isolation (unusual when considering large and widespread animals) or rapid evolution, migration, and/or replacement over a short period of time.
Of especial interest in the Los Colorados Local Fauna is the presence of protosuchid crocodylomorphs and trithelodontid cynodonts. These are the only records of these animals in the Triassic; everywhere else they suddenly appear only in the Hettangian (earliest Jurassic). It has been suggested that this indicates that this fauna extends stratigraphically over some range - from Norian or Rhaetian to Hettangian. But this is made less likely by the fact that the typically Late Triassic Riojasuchus (an ornithosuchid) is found in association with Hemiprotosuchus (a protosuchid) [Bonaparte 1970]. So either ornithosuchids really did continue through to the earliest Jurassic, or this really is a transitional fauna. For the sake of dogmatic arrangement (never a good habit) we will here go with the latter hypothesis.
The upland Thecodontosaur Community, revealed in the fissure assemblages of Cromhall, south-west England, remains diverse and little changed from the Norian; if indeed the fissure fillings from Pant-y-ffynon were (Middle or Late?) Norian. We find here again numerous Thecodontosaur small sauropodomorphs, diverse lizard-like sphenodonts, a species of Terrestrisuchus distinct in some respects from the type species (Terrestrisuchus gracilis) from Pant-y-ffynon, advanced cynodonts, early mammals, and a small herrerasaur Agnosphitys cromhallensis [Fraser et al 2002]
Similar assemblages are known from nearby regions of central Pangea. A recently discovered locality in Luxembourg, palynologically dated to Rhaetian age, shows a diverse range of animals, including Phytosaurs, Pterosaurs, possible Ornithischia, Cynodontia and Haramiyidae, Morganucodontidae and Kuehneotheriidae mammals. [Godefroit et al 1998]. And shark teeth (Synechodus rhaeticus, "Hybodus" minor, which is shown to be a Synechodontiforme, not a hybodont) and an ornithischian dinosaur tooth have been found at a new palynologically dated Rhaetian locality at Lons-le-Saunier (Jura, France). [Cuny et al 2000]
|
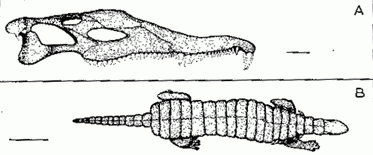
Tetrapods from the Redonda Fm: (A) Redondasaurus, (B) Redondasuchus scale bars: (A) 10 cm, (B) 20 cm
|
Aetosaur Community Further west again [Redonda Fm, New Mexico], and one finds large advanced phytosaurs (Redondasaurus - two species (R. bermanii and R. gregorii) and medium-sized aetosaurs a little like the latest Carnian Desmatosuchus, but only half the linear dimensions (Redondasuchus reseri), as well as a large unnamed aetosaur (= Paratypothorax?), the metoposaur Apachesaurus gregorii, a rauisuchian, a sphenosuchian, a procolophonid, a sphenodont, theropod dinosaurs, and a possible cynodont [Lucas 1998]. The aetosaurs are the only medium to large herbivores - a feature typical of early Norian faunas, and there are no sauropodomorphs. So either this is a conservative relict community, or it dates from an earlier period.
That the extinction of the dinosaurs and many other forms of life at the end of the Mesozoic was brought about through an extraterrestrial impact (comet or asteroid) is a popular, persuasive, but not yet universally accepted hypothesis. However a worldwide iridium layer, associated with a "fern spike" (ferns being able to regenerate in burned regions more quickly than angiosperms and gymnosperms) certainly points to some sort of worldwide catastrophe, and it has been suggested that the Chicxulub crater off Mexico provides evidence of a "smoking gun". A more comprehensive explanation is that many life forms - especially those like dinosaurs that were at the top of the food chain and so highly vulnerable to environmental fluctuations - had been highly stressed for some time by Indian vulcanism (the "Deccan Traps") and the asteroid simply executed the coup de grace. The wiping out of the dinosaurs at the same time enabled the mammals - which had at that time been flourishing albeit still small - to emerge and take over the large terrestrial niches for the first time.
There was a similar extinction event at the end of the Triassic which killed of the large Crurotarsi ("thecodonts") and allowed dinosaurs to rule as the single large terrestrial animal clade throughout the Mesozoic. While the Manicouagan in northeastern Canada is usually cited as the impact crater in this instance, it is now considered a little too early in age, and the Triassic/Jurassic (and for that matter the jurassic/Cretaceous) boundary appears to involve multiple impacts [see Asteroid/Comet Impact Craters and Mass Extinctions by Michael Paine ]. Even so, Olsen et al 2002 have recently found evidence of a K-T-type simultaneous peak in iridium traces and fern spores:
|
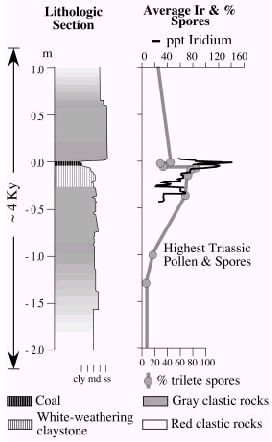
"Fine-scale correlation (left) between the Iridium anomaly and fern spike from the Jacksonwald syncline section of the Newark basin. The average Iridium anomaly is based on four localities along strike within the Newark basin, each of which have an Ir anomaly in similar positions. The duration of the interval, as based on the linear extrapolation of accumulation rates, is derived from an astronomical calibration of the entire composite Jacsonwald syncline section. [see supplemental data] Percent spore data are averaged from three sections (S. J. Fowell et al., Geol. Soc. Am. Spec. Pap. 288, 197 (1994).). Details of vertebrate assemblages and data averaging methods are given in the supplemental data. cly, claystone; md, mudstone; ss, sandstone."
text and graphic © P. E. Olsen, D. V. Kent, H.-D. Sues, C. Koeberl, H. Huber, A. Montanari, E. C. Rainforth, S. J. Fowell, M. J. Szajna, B. W. Hartline, 2002 [ref Olsen et al 2002 [Fig.2] . |
Below this iridium layer are late Triassic (Rhaetian) pollen and spores, immediately above there is the transition to typically Jurassic flora. Equally interesting is a similar transition among tetrapods, as indicated by a sudden transition in types of footprints (and hence types of animals track makers) who left them). In the following diagram, (reading from left to right under "Footprint Taxa" Eubrontes giganteus is made by a large theropod dinosaur, "Anchisauripus" a small- to medium-sized theropod, "Grallator" a small theropod like Coelophysis or Syntarsis, Batrachopus deweyii a crocodylomorph (sphenosucian?), Rhynchosauroides spp. lepidosauromorphs , new taxon B a crurotarsan, possibly crocodylomorph, Apatopus sp. by phytosaurs, and Brachychirotherium spp. by rauisuchians. It can be seen that the typically Triassic archosaurs end just below the boundary, whilst large theropods (perhaps this is Dilophosaurus) appear very soon after. Just as mammals were only able to grow large once the dinosaurs had disappeared, theropod dinosaurs, although reaching a decent medium size as early as the middle Norian, were only able to grow really big once the other big predators - in this case the phytosaurs and rauisuchians, had disappeared.
 |
"Detail of boundary section in the Jacksonwald Syncline Composite section as well as correlation to two other Newark basin boundary sections, paleomagnetic polarity data, and footprint and pollen and spore assemblage distribution. "New taxon B" is the term applied to an unnamed form by S. M. Silvestri, M. J. Szajna, New Mexico Mus. Natural Hist. Sci. Bull. 3, 439 .(1993), and cly, md, and ss, refer to claystone, mudstone, and sandstone, respectively... Interval of time represented by Jacksonwald syncline detailed section based on linear extrapolation from the average accumulation rate implied by the astronomical calibration of the Jacksonwald syncline composite section. Correlation throughout the Newark basin is based on the distinctive magnetic polarity and cycle stratigraphy and the basalts."
text and graphic © P. E. Olsen, D. V. Kent, H.-D. Sues, C. Koeberl, H. Huber, A. Montanari, E. C. Rainforth, S. J. Fowell, M. J. Szajna, B. W. Hartline, 2002 [ref Olsen et al 2002 [Supplementary Material, Fig.4]. |
Unlike the situation on land (and in keeping perhaps with the idea of several closely spaced impacts), marine fossil evidence indicates the extinction was not a single simple event. Barras 2002, studying the marine trace fossil record in England and Austria, finds that the extinction event in Austria seems to begin at an earlier stage than in England, and the fauna takes a lot longer to recover. England shows heavy bioturbation (hence, activity of many invertebrates that burrow in the mud etc) immediately prior to the Late Rhaetian extinction horizon, with  Diplocraterion,
Arenicolites, and Rhizocorallium recorded. There is then a short interval of unbioturbated sediments above the extinction horizon, indicating that for a period the seas were impoverished of life. Then strongly bioturbated sediments reappear before the boundary between the Rhaetian and the planorbis zone of the lower Hettangian (earliest Jurassic). However, Rhizocorallium does not re-appear until the upper planorbis zone (Middle Hettangian), while
Diplocraterion and Arenicolites do not re-appear until the following, upper angulata zone (latest Hettangian). Diplocraterion, on its re-appearance, is significantly smaller than its pre-event counterparts. [Barras 2002] This "Lilliputian effect" also characterizes organisms in the aftermath of the great Permian extinction [Price-Lloyd and Twitchett, 2002]
Diplocraterion,
Arenicolites, and Rhizocorallium recorded. There is then a short interval of unbioturbated sediments above the extinction horizon, indicating that for a period the seas were impoverished of life. Then strongly bioturbated sediments reappear before the boundary between the Rhaetian and the planorbis zone of the lower Hettangian (earliest Jurassic). However, Rhizocorallium does not re-appear until the upper planorbis zone (Middle Hettangian), while
Diplocraterion and Arenicolites do not re-appear until the following, upper angulata zone (latest Hettangian). Diplocraterion, on its re-appearance, is significantly smaller than its pre-event counterparts. [Barras 2002] This "Lilliputian effect" also characterizes organisms in the aftermath of the great Permian extinction [Price-Lloyd and Twitchett, 2002]
The Rhaetian sediments of Austria show a somewhat different pattern. They are thoroughly bioturbated some 60 metres below the Rhaetian/Hettangian boundary, with Rhizocorallium, Diplocraterion, and Zoophycos recorded. Immediately below the boundary itself, however, these ichnotaxa are absent. Above the boundary, laminated or layered - which means deposited with little or no stirring up of mud and sediment - sediments are common throughout the Hettangian, and here the Rhaetian trace-fossils are not recorded in until the middle Sinemurian. [Barras 2002]
Rhaetian Temnospondyls
 |
|
|
The common large Norian temnospondyl Cyclotosaurus disappears from central north Pangea in the Rhaetian, although the plagiosaurs continue; in fact they are more successful than ever, and are represented by the advanced genus Gerrothorax. A typical species of this time is Gerrothorax rhaeticus from the Rhaetic of Scania, Sweden. These large (meter long) and successful short-headed amphibians led a totally aquatic existence; in fact the presence of gills means they were unable to go ashore at all. Apart from being completely aquatic, there really wasn't much difference in appearance or habits between Plagiosaurs and the brachiopods and chigutosaurids of southern Pangea, and they were probably the direct ecological equivalent. Gerrothorax fed on fish, invertebrates, and smaller amphibians and reptiles, while themselves falling prey to the phytosaurs with which they shared their environment. Indeed, considering that they seem to have had no natural defenses against phytosaurs, Gerrothorax and other large temnospondyls did remarkably well throughout the entire period of the Late Triassic.
At the end of the Rhaetian, plagiosaurs disappear completely, whereas the brachyopids and chigutosaurids continue with little change. Perhaps conditions following the bolide impact were more severe in the northern hemisphere
Rhaetian Parareptiles
 |
|
|
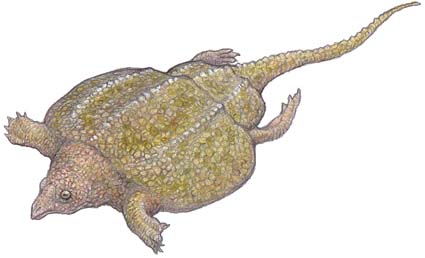
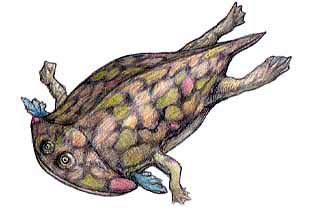
Hypsognathus was a common, successful, stocky little animal that was widespread in central west Pangea. It was the last and perhaps the most specialized of the procolophonids, with bony spines behind the skull, perhaps for protection against predators. Although the illustration shown above presents it as smooth skinned, it is just as likely to have been spiny, like the extant thorny dragon. This was probably the only defense these inoffensive animals had. Nevertheless they were remarkably successful, filling in the small herbivore niche, as they had since the earliest Triassic. Hypsognathus died out completely with the end Rhaetian extinction event, and no procolophonids are known from the Jurassic. It is not clear why these little animals, which had been so successful and widespread for so long, should completely disappear. Perhaps the bolide impact and "nuclear winter" killed off the plants on which it depended. In any case, with the disappearance of the procolophonids, the small lizard herbivore niche remained vacant for much of the Jurassic
Clade: Parareptilia
Family: Procolophonidae
Late Norian to Latest Rhaetian of Passaic Formation of New jersey, and correlative strata in Connecticut and Nova Scotia.
Terrestrial (desert/semi-desert) herbivore of the Hypsognathus community
Rhaetian Chelonians
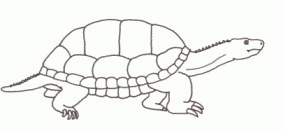
Palaeochersis talampayensisis an early turtle known from the Rhaetian Los Colorados fauna of Argentina. In the scheme of turtle evolution, Palaeochersis is the sister group to the pleurodires and cryptodires - more advanced than Proganochelys (although with a similar long tail) but, interestingly, more primitive than Proterochersis, despite the latter's earlier date. This shows how incomplete the fossil record of early turtles still is, with a number of "ghost lineages" still to be filled in.
Like the Norian Proganochelyidae, Palaeochersis was a strictly terrestrial form, as indicated by the short forelimbs. (Proganochelys quenstedti and Palaeochersis talampayensis are the only early Chelonida with forelimbs preserved) [Joyce and Gauthier 2004]. And as with Proganochelys, shared its environment quite happily with large sauropodomorphs and medium-sized aetosaurs; these three plant eaters each grazing at different levels, and hence not competing with each other.
Rhaetian Ichthyosaurs
During the Rhaetian Ichthyosaur evolution proceeded rapidly. The Parvipelvia suddenly radiated into a number of lineages (although only a few are known from fossils; the rest are "ghost lineages") following the sudden extinction of the mighty shastosaurs. Leptonectes tenuirostris, previously known as Leptopterygius, from the Rhaetic and Lower Lias of England and France, is typical of this evolutionary radiation. This intermediate grade ichthyosaur was a large marine predator appeared in the Late Rhaetian but would continue right through to the Sinemurian (Jurassic). It is the earliest known member of the paraphyletic family Leptopterygidae
Rhaetian Plesiosaurs
As well as advanced ichthyosaurs, the Rhaetian saw the radiation of an important new wave of Sauropterygia - the Plesiosauria. Thalassiodracon hawkinsi is one of the most primitive and earliest known plesiosaurs. This was a small animal, in life only about 2 to 3 meters long, known from the Rhaetic and Lower Lias of Somerset, England. It lived alongside other early representatives of the group, like Eretmosarus, which was about 3 meters long. Despite their long necks, both these animals are included in the "Pliosauria" on the basis of other features of the skull and skeleton. These animals survived into the earliest Jurassic, as part of a classic Rhaeto-Liassic marine biota. There is of course no guarantee that the poorly known stratigraphically earlier specimens belong to the same species as the later, Hettangian and Sinemurian forms
Rhaetian Placodonts
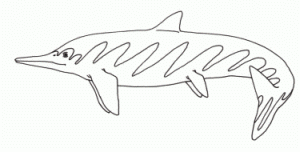
Psephoderma, which is known from the late Middle Norian through to the late Rhaetian of the Tethys ranging through to Central Laurasia (England, presumably via shallow sea), is the last of the placodonts. If its cousin Placodus was a walrus analogue, and Henodus a turtle analogue, this unusual but quite successful animal was the reptilian equivalent of the modern ray. The body is very broad and flattened. The discoidal carapace armour is in two parts, making the animal more mobile, there is a long thin tail, which does not seem to have been used in swimming. As with many other placodonts, the teeth are adapted to crushing shellfish. The main shell or carapace is made up of a large number of interlocking hexagonal osteoderms, while a smaller shell or caudal plate covers the pelvis and base of the tail. At least two species are known: the Tethyan P. alpinum and P. anglicum from the Rhaetian of England. [Naish 2000]
Body length of 1.25 meters
Links:
Psephoderma
Rhaetian Sphenodonts
Clevosaurus hudsoni was a small insectivorous sphenodontan, the remains of which are known from British fissure fills. It served as the lizard analogue of the latest Triassic; true lizards would not appear until the Middle Jurassic. This animal was more advanced than its Norian predecessors such as Clevosaurus minor, and is similar to forms found in the early Jurassic (Lower Lufeng series) of China [Fraser, 2004]. It seems to be a common Rhaetian form, and its stratigraphic range appears to be restricted to that age, which would make it a useful index fossil. However more work needs to be done before the microvertebrates of the Anglo-Welsh fissure deposits can be reliably dated.
Clevosaurus lived alongside Diphydontosaurus, a rather more primitive sphenodontian of the family "Gephyrosauridae". Without doubt, the Norian and Rhaetian fauna was very diverse, with a number of kinds of small lizard-like reptiles scurrying over rocks and hiding nooks and crannies in the late Triassic landscape.
Continued on Next Page