
| Chelonii | ||
| The Vertebrates | Protostegidae |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Anapsida ?
│
└─○Chelonii
├─Odontochelys
└─┬─Proganochelys
└─┬─Proterochersis
└─┬─Meiolanoidea
└─┬─Pleurodira
├─Paracryptodira
└─○Eucryptodira
├─○Protostegidae
│ ├─Santanachelys
│ └─┬─Bouliachelys
│ └─○"Euprotostegidae"
│ ├─Notochelone
│ └─○"Protosteginea"
│ ├─Desmatochelys
│ ├─Rhinochelys
│ └─○Protosteginae
│ ├─Chelosphargis
│ └─┬─Calcarichelys
│ └─○Protostegini
│ ├─Microstega
│ └─○Protostegina
│ ├─Protostega
│ └─Archelon
├─Plesiochelyidae
└─┬─Angolachelonia
╘═╤═Xinjiangchelyidae
╞═Sinemydidae
├─Trionychoidea
└─┬─┬─Chelonioidea
│ ├─Chelydridae
│ └─Kinosternoidea
└─Testudinoidea
|
Contents
Overview |
Taxa on This Page
|
|
| Archelon ischyros, Early Campanian of the Cretaceous Inland Sea, life reconstruction by , GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
The Protostegids were the first ocean going testudines, and also included some of the largest. They were highly convergent with leatherback turtles (Dermochelyidae), and original morphological cladistic studies grouped them together in the superfamily Dermochelyoidea (Gaffney & Meylan 1988) or clade Pandermochelys (Joyce et al 2004). Subsequent computational-cladistic analysis including the ancestral protostegid Santanachelys gaffneyi shows that the Protostegids are not Cheloniodea at all, but belong to more ancient and primitive lineage of stem-cryptodires or even stem testudines dating back to the late Jurassic, and that the similarities to Dermochelyids are the result of convergence. As Joyce notes:
" representatives of the Protostegidae are markedly primitive in many of their characteristics, including the absence of formed cervical articulations and the presence of elongate first thoracic ribs. A placement of Protostegidae within Chelonioidea thus either requires the systematic reacquisition of these primitive traits within Chelonioidea, or the independent loss of these traits in numerous cryptodiran lineages." - Joyce (2007) p.66
 |
| Size comparison of three members of the Protostegidae family : Protostega gigas, Archelon ischyros, and Notochelone costata, with a human for scale. By TheF, silouettes from artwork by by Dmitry Bogdanov. GNU Free Documentation/Creative Commons Attribution license, via Wikimedia |
As with other marine tetrapods the tendency towards large size was prominent (right). Some such as Bouliachelys, Desmatochelys, and Protostega equalled or exceeded the extant leatherback in size, while Cratochelone and Archelon would have had shells upto 3 meters in length and flipper spans of upto 5 meters. These animals would have made an impressive sight, swimming among the other equally huge reptiles of the Cretaceous seas.
Like many latest Cretaceous animals, protostegids did not make it to the end of the Mesozoic (ref Parham & Pyenson 2010)). After flourishing for some fifty million years they disappeared during the mid-Campanian, some ten million years before the asteroid strike and K-T extinction. During the very latest Cretaceous (Late Campanian and Maastrichtian) they were replaced by new dynasties of sea turtles, some of which equalled them in size. By the early Cenozoic the two lineages of modern sea turtles (Cheloniidae and Dermacheylidae) were ecologically diverse and well established (Mlynarski 1976, Parham & Pyenson 2010) MAK130313
 |
| Replica of a 75 million year old Archelon ischyros from South Dakota, now hanging out in the Wallace Building on the University of Manitoba campus, Winnipeg, Manitoba Canada. Posted to Flickr as Super Arky, photo Mike Beauregard. Creative Commons Attribution 2.0 Generic license, via Wikipedia |
Range: late Aptian-early Albian to early-mid Campanian, worldwide
Phylogeny: Eucryptodira : Plesiochelyidae + Thalassemyidae + (Angolachelonia + Xinjiangchelyid - Crown Cryptodire clade) + * : Santanachelys + (Bouliachelys + "Euprotostegidae")
Characters: "Nasal present; jugal–quadrate contact, excluding quatrado–jugal from lower cheek margin; lower cheek emargination absent; vomer–palatine contact lost, palatines medially meeting; foramen palatinum posterius open postero–laterally; pterygoid extending onto mandibular articular surface of quadrate; foramen posterius canalis carotici interni between pterygoid and basisphenoid; rodlike rostrum basisphenoidale; third (or second) cervical centrum biconvex; lateral process of pubis extending anteriorly beyond mideal portion of pubis; lateral process of humerus enlarged within the anterior portion of shaft, not easily visible from ventral view; radius with middle portion bent toward anterior; cervical scute overlying more than half the width of nuchal; thick neurals with median keel." - Hirayama 1998 p.707.
Further description: Carapace represented merely by a row of marginals, but the plastron very strongly developed and composed of thick ossifications. Skull with descending parietal plates. Humerus with the radial process short, blunt, and approximated to the head.Zittel, Eastman, et al (1902) pp.197
Comments: Edited from Wikipedia: As the earliest marine turtles, the protostegids set the general body plan for future lineages. They had a generally flattened body, with long oar-like front appendages specially evolved for swimming in the open ocean. Convergent with the Dermochelyidae, with which they probably shared a very similar ligfestyle. protostegids possessed extremely reduced carapaces. Some specimens had skeletal protrusions from their ribs almost wrapping around the turtles' bodies in place of a complete shell. They also had somewhat reduced plastrons, although not to the extent of dermochelyids. As with modern sea turtles, protostegids had sharp beaks. One of the defining characteristics of the members of the family was their almost-disproportionately large heads. Specifically, some specimens of Archelon have been found with heads a meter long.
Analysis of fossil organs has revealed entire stomachs containing fossilized shellfish (Hirayama 1998). In turn, they were preyed upon by the apex marine predators of the time. Fossil protostegids have been found with tooth impressions from the large lamnid sharks (Shimada et al 2002). Two specimens of Protostega gigas have been discovered to have tooth marks from large sharks. In addition, teeth of the extinct shark Cretoxyrhina mantelli (a Cretaceous equivalent of the Great White) have been found embedded in at least one Protostega skeleton.(Shimada & Hooks 2004) - Wikipedia
Classification and phylogeny:
In 1888, the Belgian zoologist George Albert Boulenger published his classification of the Testudinata within the 9th edition of the Encyclopaedia Britannica. The genus Protostega was placed within the family Sphargidae (= Dermochelyidae) under the suborder Athecae. A year or so later, the entire suborder was downgraded by Karl Alfred von Zittel into a family within the Cryptodira. (Baur 1890)
Original cladistic studies supported the relationship of Protostegidae and Dermochelyidae, the two being considered monophyletic families and sister taxa (Gaffney & Meylan 1988, Hirayama 1998, Gaffney & Meylan 1988, Meylan & Hirayama 2000), the two teogether being included in the clade Dermochelyoide
In 1994, Hirayama proposed a three-family subdivision of the sea turtle superfamily based on cladistic analysis; Protostegidae was given full, formal family status in the system, containing most of the extinct genera, including Archelon, and a previously undescribed protostegid. (Hirayama 1994] The unidentified specimen was fully described in 1998, as Santanachelys gaffneyi. It was appended to the family after the new species was described. This specimen was later to be analyzed to be the family's oldest member.[Hirayama 1998]
But computational phylogenetic analyses conducted by Joyce (2007) and later workers, which included the most primitive protostegid species, Santanachelys gaffneyi, recovered the family as unrelated primitive turtles that only resemble leatherback turtles by convergence, although precise details differ. Joyce (2007) recovered Protostegidae as stem eucryptodires outside the Cryptodira crown group. Solnhofia parsonsi was found to be closely related. Anquetin, 2009 and 2012 arrived at a similar result, although he emphasised that future studies should include more protostegids to confirm this phylogenetic placement. Using a more restrictive dataset and a newly discovered African species, Mateus et al (2009) found Thalassemys marina to be the sister taxon of Santanachelys. Sterli, (2010), Sterli & de la Fuente (2011), Pérez-García 2012, recovered Santanachelys (and by implication the rest of the Protostegidae) as stem testudines outside the crown group of turtles (the least inclusive clade containing cryptodirans and pleurodirans). These results confirm that protostegids represent an independent lineage of marine turtles originating from primitive Late Jurassic forms, but astonishingly convergent with latest Cretaceous and Cenozoic Chelonoidea (Wikipedia modified by MAK130307)
Links: Oceans of Kansas, Wikipedia, Geologic History of Sea Turtles (short notes), Mikko's phylogeny (dendrogram only) MAK130313
 |
| Life restoration of Santanachelys gaffneyi. by Smokeybjb, GNU Free Documentation/Creative Commons Attribution license, via via Wikipedia |
Horizon: Santana Formation (late Aptian - early Albian) of North-East Brazil
Phylogeny: Protostegidae : (Bouliachelys + "Euprotostegidae") + *
Comments: Santanachelys gaffneyi is the earliest known sea turtle. This 20-centimeter long animal resembled a terrapin in many respects, its marine lifestyle is indicated by the enormous salt glands around the eyes. This is confirmed by the fossil remains being found in marine sediments.
The kidneys of modern marine turtles are unable to deal with the excess salt which would result in dehydration from eating salty food or drinking seawater. To solve this problem they have enormous tear (lachrymal) glands, each bigger than their brain, adapted to excrete a salt solution more concentrated than seawater. Their presence is evident in fossil forms as well by of the large foramen in the skull where salt glands would be. From this we know that Santanachelys had the same type of huge salt glands situated near its eyes which allowed it to drink saltwater without dehydration.
Yet despite this advanced feature, and other characteristics shared with of modern sea turtles, it retained a number of primitive characteristics. For example, the metacarpals and short digits of the feet were still were distinct and movable, as in non-marine turtles, unlike the fully fused and rigid, hydrodynamic paddles that advanced sea turtles possess. This shows that the evolutionary return to oceanic waters of the sea turtle line evolved before the finalization and streamlining of the paddles. (Hirayama 1998)
 |
| Santanachelys gaffneyi from Hirayama 1998 |
For several years Santanachelys gaffneyi was considered the earliest known Pandermochelyian as well as the oldest panchelonioid (Joyce et al 2004 p.1000). More recent cladistic analysis showed it to be a much more primitive form, totally unrelated to conventional sea turtles. This means that the many apomorphies and similarities between protostegids and dermochelyid chelonoidians (including - Hirayama 1998) are the result of convergence (homoplasy) rather than shared ancestry (homology). Hence identical solutions, such as for example giant tear glands for processing salt and preventing dehydration - evolved completely independently from only very distrantly related ancestors as a way of coping with a particular environmental problem. This sort of astonishing convergence means that one of the central pillars of cladistics - parsimony - is an unreliable assumption. One may even wonder to what extent such parallelism would feature in extraterrestrial life; would alien biota, rather than resembling the buf eyed mosnters of B-grade movies, actually be strongly convergent with terran life-forms? (as argued by paleontologist Simon Conway Morris (Conway Morris (year))) MAK130313
Links: Wikipedia, Studio d'Arte Corvo
Range: Early Cretaceous (Late Albian) of Australia
Phylogeny: Protostegidae : Santanachelys + ("Euprotostegidae" + * )
Comments: this primitive protostegid is similar to Santanachelys in having a poorly developed secondary palate, which shows that this feature evolved convergently in advanced protostegids and cheloniids. The hooked beak (an adaptation for feeding on pelagic ammonites; Hirayama 1994, 1997) of Bouliachelys is ismilar to that of more derived giant forms like Protostega, and is another convergent adaptation. Bouliachelys is the largest known before the Late Cretaceous - shows that early sea turtles were both larger and more diverse than was previously thought, The skull of Bouliachelys suggests an animal some 50% larger than Notochelone (maximum body length less than 1 m; Molnar 1991); an even bigger but very poorly known sympatric taxon, Cratochelone may have had a body length approaching 4 m, which puts it in the Archelon range. (Kear & Lee 2006)
Range: Late Albian to Mid Campanian
Phylogeny: Protostegidae : Santanachelys + (Bouliachelys + * : Notochelone + "Protosteginea" )
Characters: (Hirayama 1998)
Comments: "Euprotostegidae" is here used as in informal name for the group containing Notochelone and higher protostegids
 |
| Notochelone costata, an early protostegid turtle from Early Cretaceous (Albian) of Australia. Artwork by Dmitry Bogdanov, GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
Range: Late Albian of Australia
Phylogeny: "Euprotostegidae" : "Protosteginea" + *
Comments: A medium-sized sea turtle, about the same size as the modern green turtle. It is thought that these animals fed on benthic molluscs. (durophagous) (Wikipedia)
Range: Turonian to Mid Campanian
Phylogeny: "Euprotostegidae" : Notochelone + * : Desmochelys + Rhinochelys + Protosteginae
Characters: Foramen palatinum posterius absent; pterygoid narrow, C-shaped; large processus trochlearis oticum involving deep notch of quadrate; first suprapygal absent. (Hirayama 1998) medial contact between palatines; foramen posterius canalis carotici interni located between the basisphenoid and pterygoid; coracoid longer than scapula; and radius bends posteriorly Hooks 1998
Comments: "Protosteginea" is here used as in informal name for the group containing Desmatochelys, Rhinochelys, Cimochelys, "Protostega" eaglefordensis and the protostegids (= protosteginae)
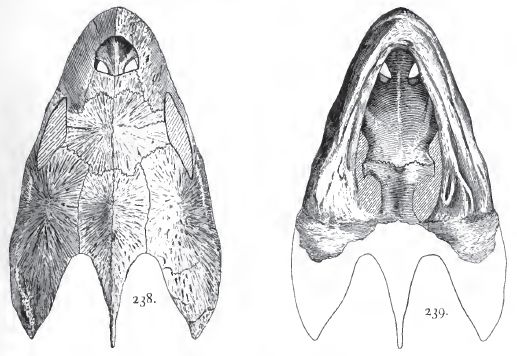 |
| Desmatochelys lowi. Skull of type, length 20.5 cm, width 14.5 cm. Niobrara formation (Santonian), Kansas. From Hay (1908) p.185, figs 238 and 239; public domain (copyright expired). |
Range: Turonian to Santonian-Campanian of North America
Phylogeny: "Protosteginea" : Rhinochelys + Protosteginae + *
Descriptions: Skull with temporal region roofed over as far backward as the occipital condyle. Large nasal bones present. Choanae placed well forward, not underfloored by the vomer, maxillae, and palatines. Small posterior palatine foramina present. Humerus indicating a paddle-like fore limb. Plastron loosely joined to the carapace.- Hay (1908) p 184
Comments: Not to be confused with Dermochelys (although the two are similar in size and probably habits), this was a large intermediate grade protostegid, with a plastron length of about 1.5 meters (Mlynarski 1976 pp.50-51). Made the type for its own family Desmatochelyidae. The poorly known genera Atlantochelys and Neptunochelys were provisionally included (Hay (1908) p 184) but are more likely incertae sedis, perhaps synonyms of better known or more common species. For some time of uncertain evolutionary relationship, it has been placed in the Cheloniidae incerta sedis (Mlynarski 1976) and with Corsochelys as the subfamily Desmatochelyinae in the family Dermochelyidae. Cladistic analysis shows it to be a member of the Protostegidae (Hirayama 1998, Kear & Lee 2006), and it is sometimes grouped with Rhinochelys, a much smaller animal of a similar grade. As well as being known from (wherever) fossil remains of the genus, have been found in the Trent River Formation (Santonian-Campanian) of the Comox Basin of eastern Vancouver Island (Nicholls 1992), which extends the stratigraphic and geographic range extensions for the genus Desmatochelys to the Pacific coast . It is quitre likely that, like modern sea turtles, these animals had an ocean-wide distribution.
 |
| Rhinochelys nammourensis, Cenomanian of Lebanon. Specimens like this showed the synonymy of Rhinochelys (known only from the skull) and Cimochelys (known only from the shell). Photo by Ghedoghedo, GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
Range: Cenomanian of England, France, and Lebanon
Phylogeny: "Protosteginea" : Desmochelys + Protosteginae + *
Comments: The following is edited and adapted from a Google translation of the French Wikipedia
Rhinochelys is known mainly from fossil skulls found in the early Late Cretaceous (Cenomanian) of England and France in the 19th century and described by Owen, Seeley, and Lydekker. In all, more than twenty species have been named and described, mostly by Seeley in 1889. The genus was revised by Collins in 1960 (Collins, 1960), who recognised only three English species - the type species Rhinochelys pulchriceps ( Owen, 1851), R. cantabrigiensis Lydekker, 1889, R. elegans Lydekker , 1889, and the French R. amaberti Moret, 1935. The rest are ether synonyms or scrappy. The skulls are from rather small sea turtles, generally 4 to 6 cm in length (Mlynarski 1976 p.39) Collins also mentions Cimochelys benstedi , known only by its shell, but found in the same strata as Rhinochelys, although it is possible these two genera are the same. Based on a diagnosis of post-cranial characters, Collins included Cimochelys benstedi among Protostegidae, along with Chelosphargis.
The identity of Cimochelys and Rhinochelys was accepted by Hirayama in his revision of the family Protostegidae in 1994 (Hirayama, 1994). He also synonymises Collins' four species under the name Rhinochelys pulchriceps , to which he adds Rhinochelys benstedi (the former Cimochelys).
In 2006, Tong et al 2006 described specimens from Lagerstätte of Nammoura at Lebanon as a new species Rhinochelys nammourensis . The fossils, very well preserved and nearly complete, undoubtedly belong to Rhinochelys given their cranial characters, although they differ enough to justify placing them in a new species. The shape of the carapace, also very similar to that of Cimochelys, confirming that both genera are the same. - Wikipedia
Given the identity of Cimochelys and Rhinochelys, the present author (MAK) fails to see the reason for retaining R. benstedi as a separate species. That leaves only two species, Rhinochelys pulchriceps ( Owen , 1851), and Rhinochelys nammourensis Tong et al 2006.
Mlynarski 1976 pp.38-9) places Rhinochelys with Calcarichelys and Chelosphargis in Zangerl, 1953a's paraphyletic (Hooks 1998) subfamily Chelospharginae MAK130313
Links: French Wikipedia, Mikko's Phylogeny Archive (retains four species)Protosteginae : Archelon, Calcarichelys, Chelosphargis, Microstega, and Protostega
Range: Coniacian to Mid Campanian
Phylogeny: "Protosteginea" : Desmochelys + Rhinochelys + * : Chelosphargis + (Calcarichelys + Protostegini)
Characters: Prefrontal-postorbital contact present; scar for M. latissimus dorsi and teres major located at middle of shaft; star-shaped, hyo–hypoplastra. (Hirayama 1998); absence of the foramen palatinum posterius; a basioccipital with knob-like processes fitting into sockets on the posterior end of the pterygoids; the presence of only one suprapygal; and a T-shaped entoplastron that is not sutured to the other plastral elements - Hooks 1998 (as "Protostegidae")
Comments: As evidenced from theire respective character and genus lists, Hirayama and Hooks, writing in the same year, define the same clade using different synapomorphies, and in Hooks' case two extra genera, Calcarichelys and Microstega. Hooks refers to this clade under the name Protostegidae, but as that family ranking is now universally applied to a more diverse range of taxa we have referred to it as the subfamily Protosteginae. MAK130313
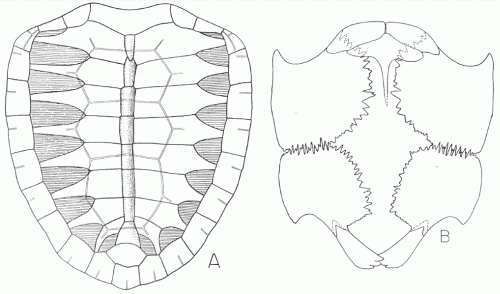 |
| Chelosphargis advena, No complete specimens are known; this is a reconstruction of the carapace (left) and plastron (right) from available materials. The left side represents the juvenile shell, the right the adult. From Zangerl, 1953a fig 26, p.87 |
Range: Coniacian/Santonian to Early Campanian of North America
Horizon Niobrara Chalk, Kansas and Mooreville Chalk, Alabama
Phylogeny: Protosteginae : (Calcarichelys + Protostegina) + *
Description: " Primitive genus of Protosteginae of small or medium size. Skull with blunt, straight premaxillary beak. Frontal bones large and with lateral processes toward the orbital rims. Prefrontals excluded from sagittal contact by nasal bones. Otic and exoccipital area very similar to condition in cheloniid turtles. Lower jaw with long symphysis; rami fused, even in juvenile specimens. A slight, but sharp sagittal crest on triturating surface of lower jaw. Carapace laterally with fontanelles of moderate size in juvenile specimens. Neural bones forming blunt, low keel in midline. Peripheral edge of carapace even. Pygal plate entirely separating eleventh pair of marginals." - Zanger 1953a p.80
Comments: The most primitive of the Protostegines, Primitive genus of Protosteginae of small or medium size. The species is quite common, but is represented mostly by very young individuals. Formerly Protostega advena Hay (1908); the over-all appearance and the major proportions of the skull of this species are very similar to those of Protostega. Zangerl, 1953agroups Calcarichelys and Chelosphargis in a new subfamily Chelospharginae. Cladistic analysis by Hooks 1998 shows the latter to be a paraphyletic taxon, which makes it invalid as a cladistic taxon but not as a Linnaean or Evolutionary taxon. MAK130313
Calcarichelys gemma Zanger 1953
Range: Santonian to Early Campanian of North America
Horizon: Mooreville Chalk, Alabama; and Niobrara Chalk, Kansas.
Phylogeny: Protosteginae : Chelosphargis + (Protostegini + * )
Characters: a small specialized protostegid closely allied to Chelosphargis, but with uniquely specialized neural keel and strongly serrate peripheral margin except in front). Unusually high, sharply pointed neural plates alternate with nearly flat saddle-shaped ones. Last keel-thorn located on suprapygal. Pygal plate very narrow. Hypoplastron nearly rectangular. - Zanger 1953a p.119, 129
Comments: This small (shell about 30 cm) but bizarre protostegid was equipped with a row of spines along the dorsal mnidline of the shell, alternating with more typically smooth plates. The spines doubtless were defence against predators, as unlike many other protstegids this little animal couldn't rely on sheer size. It was first discovered Burkville near Montgomery in Alabama, by CM Barber in 1946 and described in 1953 by Rainer Zangerl. Zanger's diagnosis of Calcarichelys gemma is based on the study of another fossil discovered in 1949 by Barber Greene, which was particularly complete and well preserved, nicknamed "the rock" by its discoverer, and which is at the origin of name of the species. (French Wikipedia) . Along with the closely related Chelosphargis it has been found in both the Niobrara Chalk of Kansas and in the Mooreville Chalk of Alabama, although unlike Chelosphargis it is best know from the latter locality. Zanger 1953a includes the two monospecific genera in a new subfamily Chelospharginae, representing a more primitive level of protostegid organization, although with certain specialized features paralleling those in the Protosteginae (= Protostegini) (Zanger 1953a p.131). It is interesting that these smaller and more morphological primitive animals ranged widely from the inland sea to the open Atlantic, whereas the giant and highly derived protostegids are so far known only from the inland sea, although there is no reason why they could not have had a wider distribution. So far, no protostegines have been found beyond North America, indicating either a lack of suitable starta, or that these great turtles were limited in distribution to the Cretaceous inland sea of mid-west North America. Protosphargis is a convergent but unrelated form from the Latest Cretaceous of Europe. MAK130401
Range: Santonian to Mid Campanian of North America
Phylogeny: Protosteginae : Chelosphargis + (Calcarichelys + * : Microstega + Protostegina
Comment: The "higher" protostegines. As with Hominidae/inae/ini/ina this taxon's suffic is based on the cladistic sequence of branching, rather than Linnaean tribes which although having the same suffix pertain to a different taxonomic system
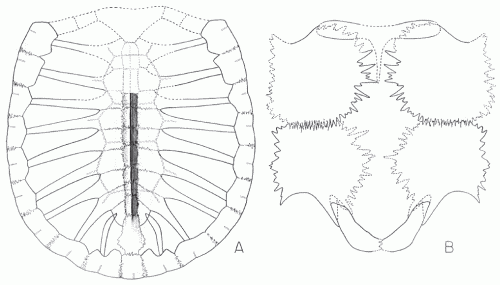 |
| Microstega copei, carapace (left) and plastron (right). From Zangerl, 1953a fig 20, p.79 |
Microstega copei (Wieland 1909)
Synonyms: Archelon copei, Protostega copei
Range: Santonian or Early Campanian of North America
Phylogeny: Protostegini : Protostegina + *
Comments: Based on a disarticulated skeleton of moderate size (YPM 1787), this species was described by Wieland, 1909 as a new species of Protostega. Zangerl, 1953a noted the even keel formed by the neural series of bones in the carapace is similar to that of Archelon ischyros, unlike the uneven, interrupted keel of Protostega. He therefore transferred the species to Archelon, as a more primitive species of that genus.. Hooks 1998 using cladistic analysis discovered this species is more basal than both Archelon ischyros and Protostega gigas.and gave it its own genus as well as making it sister taxon of the Protostegina. As there is very little morphological difference between Protostega and Archelon it probably really doesn't matter what genus this species is allocated to MAK130315
| Protostega gigas. Photo by Claire H. from New York City, GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
Range: Santonian to Mid Campanian of North America
Phylogeny: Protostegini : Microstega + * : Protostega gigas + Archelon (= Protostega ischyros)
Description: "Marine turtles with the fore limbs converted into flippers resembling those of the Cheloniidae. Carapace greatly reduced, the disk extending not one-half the distance toward the distal ends of the ribs. Peripherals present. Plastron loosely connected with the carapace and with a large median fontanel. Entoplastron T-shaped, with the lateral wings elongated and distally expanded. Epiplastra not certainly known. Xiphiplastra short and bent. Skull large, temporal region broadly roofed over. Region in front of the orbits elongated. Jaws with large crushing-surfaces. Choanae far forward; not underfloored by the surrounding bones." - Hay (1908) p.189
Comments: The above is Hay's description of Protostegidae sensu Protostega + Archelon. These were the last and largest of the Protostegids Zangerl, 1953a makes this the subfamily Protosteginae (Mlynarski 1976 pp.39-40), while Hooks 1998, using cladistic analysis, gives this clade the Linnaean rank of subtribe as Protostegina. But in asmuch as Protostega and Archelon represent a morphological and stratigraphic sequence, only attachment to the coolness factor associated with the name Archelon, and the cladistic phylogenetic reluctance to attribute more than one species to each genus, prevents Archelon from becoming a junior synonym of Protostega, as Protostega ischyros (Wieland), and in fact Williston (1897) did use this binomial, although it seems that no-one followed him in this. Hooks (1998) placed the contemporary Archelon copei in a new, somewhat more basal genus, Microstega (Hooks 1998), although ll three species are probably similar enough to be considered congeneric. MAK130313
| Protostega gigas, life reconstruction by Dmitry Bogdanov, GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
Horizon: Type specimen: Smoky Hill Chalk of western Kansas (Santonian)
Phylogeny: Protostegina : Archelon (= Protostega ischyros) + *
Description: Descending parietal plates well developed. Carapace intermediate between Dermochelyidae and Chelonidae, with several primitive characters regarded as ancestral to both. Plastron with larger bones and a much smaller fontanelle than in Dermochelys. Body elongate, the posterior end truncated; limbs paddle-shaped. Post-axial border of humerus more orhis genus less deeply emarginate. (Zittel, Eastman, et al (1902) pp.198). Premaxillary beak less developed than in Archelon. Maxilla with a rather broad grinding surface, which extends backward to behind front of orbit. Lower jaw with the rami early co-ossified. Entoplastron T-shaped, with the middle third of the anterior border concave from side to side, the distal ends convex. Radial process of humerus large. Hay (1908) p.190,
Comments: Differs from Archelon only in somewhat smaller size and earlier stratigraphic horizon, this is clearly an ancestral species top the latter. Average plastron length of 1.2 meters (Mlynarski 1976 p.39), maximum overall length upto 3 m. Like the similar-sized modern leatherback sea turtle, with an overall length of upto 2.7 m, Protostega's carapace lacked scutes, making it weaker but also lighter. (Wikipedia). Protostega most likely fed on plegaic molluscs such as belemnites
A number of species have been attributed to to this genus. Protostega copei is a valid taxon from the Niobora of Kansas and has been given a distinct generic name by Hooks 1998; see Microstega. Protostega dixie from Selma Formation (Mooreville Chalk), of Alabama is considered a distinct species by Zangerl, 1953a but a synonym of Protostega gigas by Hooks 1998. Protostega potens Hay is based on a partial and weathered skeleton (AMNH 180), also from the Niobora formation. Zangerl, 1953a considers it a distinctive species but Hooks 1998 makes it a junior synonyms of Protostega gigas. The fossil indicates a turtle larger than any known specimen of P. gigas, but considerably smaller than the giant type specimen of Archelon ischyros. It may simily be a very large P. gigas. "Protostega" eaglefordensis is a large but poorly known protostegid known from partial remains from the Eagle Ford Formation, Texas. The limbs and girdles are more primitive than in any known large protostegid Zangerl, 1953a p.89. The phylogenetic relationships are unresolved but according to (Hooks 1998) it does not belong in the genus Protostega. MAK130313
Links: Oceans of Kansas, Wikipedia (stubby article but comprehensive bibliography).
 |
| Type specimen (YPM 3000) of Archelon ischyros in the Yale Peabody Museum, Yale University. Amazingly enough, this giant animal's skeleton represents by no means the largest known Archelon. From Frederic A. Lucas, 1902, Animals of the past, copyright expired , public domain, via Wikipedia |
Archelon ischyros Wieland. 1896
Range: Upper Pierre shale (Mid Campanian) of South Dakota and Wyoming.
Phylogeny: Protostegina : Protostega + *
Description: Premaxillary beak strongly developed. Crushing-surface of upper jaw mostly on the premaxillaries; that on the maxillae extending back only to opposite the choanae. Lower jaw with the rami not co-ossified at symphysis; at least, not until old age. Entoplastron T-shaped, with the anterior border concave from end to end. Radial process of humerus feeble. Hay (1908) p 202
Comments: The appropriately named Archelon ("ruler") is with the poorly known Gigantatypus the largest sea turtle that has ever been described, although the freshwater Stupendemys may have grown even larger. The type specimen of Archelon (YPM 3000) was collected from the Pierre Shale of South Dakota by Dr. G.R. Wieland in 1895 and described by him the following year (Wieland, 1896) This and other specimens were recovered near the South Fork of Cheyenne River in South Dakota, in the upper Pierre formation, below the Judith River formation (Mid Campanian). From the length of the neck and the carapace Wieland estimates that the total length of type specimen was about 3.5 meters. The estimated total length of the carapace of the original animal is about 1.9 meters. The width, exclusive of the peripherals, must have been about 2 meters; including the peripherals, about 2.5 meters. Hay (1908) pp 202-203. Wieland also described another, more fragmentary specimen of similar size from the same formation and locality, which he named Archelon marshii, and which is clearly a synonym of A. ischyros
An even larger Archelon fossil was found in the Pierre Shale of South Dakota in the 1970s, is more than 4 metres long, and about 4.9 metres wide from flipper to flipper. Since only the tiniest fraction of organisms that ever lived are fossilised, and given that some recent sea turtle specimens are of larger than average size, it is similarily very likely that there were even larger Archelons, in keeping with exaggerated measurements in non-technical literature of flipper to flipper span of 5.5 meters.
 |
| Archelon ischyros, mounted cast, closeup of head, Canadian Museum of Nature, Ottawa, Ontario, Canada. The upper jaw's very large beak is evident Photo D. Gordon E. Robertson, Creative Commons Attribution license, via Wikipedia |
Apart from larger size, Archelon is anatomically very similar to its predecessor Protostega gigas, and Williston 1897 includes it in the same genus; a reasonable approach that is not acted on because of the romantic trendency to make genera of large impressive animals monospecific (Baronyx and Suchimimus are two other examples of genera that are only distinct at the species level)
.Unlike most turtles, Protostega and Archelon did not have a solid shell, but instead had a skeletal framework supporting a leathery or bony carapace. In these and other respects they are convergent with Cenozoic and modern Dermochelyds or leatherback turtles. Other distinguishing features include a pointed tail, a narrow skull, a relatively narrow, high-vaulted shell, and a pronounced overbite. The live weight of an Archelon ischyros is estimated at more than 2,200 kg. The bite was probably very strong, optimized for feeding on pelagic mollusks such as squid and belemites. The specimen exhibited by the Museum of Natural History in Vienna is estimated to have lived to be a century old (Wikipedia).
The head alone was enormous, upto a meter long, and the formidable beak, with its broad crushing-surface, indicates a diet of mollusks and crustaceans. Associated with the fossil were various genera of belemnite tetrabranchiate cephalopods, and the jaws of this giant turtle seem to have been well adapted to pick up and crush such objects. Hay (1908) p.208,
Archelon's beak more strongly developed than in Protostega Hay (1908) p 202), presumably as the result of allometric growth (like large deer which grow proportionally larger antlers with lareger body size).
MAK130313
{kind=link}
{kind=link}
{kind=link}
{kind=link}