
| Chelonii | ||
| The Vertebrates | Trionychoidea |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Anapsida ?
│
└─○Chelonii
├─Odontochelys
└─┬─Proganochelys
└─┬─Proterochersis
└─┬─Pleurodira
├─Paracryptodira
└─┬─Protostegidae
╘═╤═Xinjiangchelyidae
└─○Centrocryptodira
╞═Sinemydidae
├─┬─○Trionychoidea
│ ├─Basilochelys
│ └─┬─○Adocusia
│ │ ├─Yehguia
│ │ └─┬─○Adocidae
│ │ │ ├─Adocus
│ │ │ └─Shachemydinae
│ │ └─○Nanhsiungchelyidae
│ │ ├─Kharakhutulia
│ │ └─┬─Hanbogdemys
│ │ ├─○"Nanhsiungchelyinae"
│ │ │ ├─Anomalochelys
│ │ │ └─Nanhsiungchelys
│ │ ├─Zangerlia
│ │ └─Basilemys
│ └─┬─Peltochelys
│ └─○Trionychia
│ ├─Carettochelyidae
│ └─○Trionychidae
│ ├─┬─Plastomeninae
│ │ └─Cyclanorbinae
│ └─Trionychinae
└─○Durocryptodira
├─┬─Chelonioidea
│ └─┬─Chelydridae
│ └─Kinosternoidea
└─Testudinoidea
|
Contents
Overview |
Taxa on This Page
|
| The African or Nile softshell turtle, Trionyx triunguis (Forskål, 1775), a modern representative of this most ancient of cryptodiran lineages. Photo by Dan Schwartz, Creative Commons Attribution license, via Wikipedia |
The Trionychoidea are phylogenetically the most controversial turtle taxon, since molecular phylogeny has them diverging very early from the rest of the cryptrodires, whereas some morphological analyses place them next to the Kinosternoidea.
Stratigraphically, they are oldest extant cryptodire clade. An extensive Mesozoic record includes the Cretaceous genera Adocus, Basilemys, Nanhsiungchelys, and Peltochelys, (Joyce et al 2004, p.1002), and Yehguia tatsuensis, late Jurassic (Oxfordian) of China (Danilov & Parham, 2006). The early Cretaceous Sandownia harrisi is sometimes included here (Meylan et al 2000), although it could equally belong to a distinct and more primitive eucryptodire lineage, the Angolachelonia (Mateus et al 2009)
Molecular evidence also supports the very early appearance of Trionychia (Shaffer, 2009), which either branched off very early from the rest of the Cryptodira, or are a paraphyletic group from which other extant cryptodires developed (Krenz et al 2005). In any case it seems that pleurodires, trionychoids, and non-trionychid cryptodires (Durocryptodira Danilov & Parham, 2006) represent a trichotomy that emerged during the mid Jurassic (Krenz et al 2005 p.189). Yet if the Trionychoidea and Kinosternoidea are distinct taxa that deverged early in eucryptodire history, the number of morphological convergences required that would consistently place them together in any cladistic parsimony analysis is simply incredible (although no worse than other moleciular phylogenies when considered morphologically, e.g. Squamata, Artiodactyls, etc). Also remarkable would be the absense of primitive characteristics in extant forms (in contrast to the Protostegidae which while converging on Chelionoidea also have a number of primitive features). On the other hand, the absense of any non-trionychoidan crown cryptodire clades prior to the Latest Cretaceouys is also remarkable. If this morphological interpretation is correct than the cryptodires have so many Jurassic-to-Mid Cretaceous ghost lineages that they seem as Cretaceous whales or rodents. Another alternative, which no-one has apparently suggested, is that the diverse and abundant Mesozoic clade Adocusia are not stem-Trionychia at all, but convergent due to similar lifestyle, in the manner of protostegids and dermocheylids. For now we have gone with the molecular and stratigraphic hypothesis over the morphology-only one. MAK130302
 |
| Chinese softshell turtle Pelodiscus sinensis. Photo from here, via CKT. |
Trionychoidea Gray, 1825 sensu Mlynarski, 1976
Synonyms: Trionychidea Bell, 1828, Trionychida Gray, 1870, Trionychoidae Meylan & Gaffney 1989, Pantrionychia Joyce et al 2004
Range: late Jurassic (Oxfordian) to Recent
Phylogeny: Centrocryptodira : Durocryptodira + * : Basilochelys + (Adocusia + (Peltochelys + Trionychia))
Comments: "Trionychoidea" is used here instead of "Trionychia" because it includes extinct as well as crown taxa, and instead of "Trionychoidae" to conform to the standard Linnaean suffix for superfamily. The Trionychoidea are a well defined and specialised assemblage of cryptodire turtles that first appear in the fossil record in the late Jurassic, they are by many tens of millions of years the oldest lineage of extant cryptodires, by the late Cretaceous they were a ubiquitious element in Laurasian freshwater faunas. Among the many lineages from this time are the Nanhsiungchelyidae, Adocidae, Carettochelyidae, and Trionychidae, along with a number of monogeneric groups. The Nanhsiungchelyids are particularily intersting as they independently evolved as large terrestrial forms, equivalent to Cenozoic testudinids.
Hennigian morphological analysis places them as a specialised clade, e.g. Gaffney, 1975, Meylan 2001, Gaffney & Meylan 1988, Meylan and Gaffney (1989), and Meylan 2001, on the basis of the following characters: Small stapedial artery, with large palatine artery and canalis caroticus lateralis; significant contribution of palatine to side-wall of braincase; tenth body vertebra free from costals; thelial process present; maxillary 'tooth' present; caudifibularis muscle present (Gaffney & Meylan 1988 via CKT110603). Gaffney and Meylan propose the phylogeny Centrocryptodira : + * : Kinosternoidea + Trionychoidae. More recent computational-statistical phylogenetic found much less morphological support for this clade; according to Joyce 2007 p.69, the clade Trionychoidea is only weakly supported in this analysis by a single character, listed above, the contribution of the palatine to the lateral brain case wall. He found the additional characters to be either problematic or the result of homoplasy. Molecular phylogeny goes further and has unanimously fails to support its monophyly. Beginning in the late 1990s, molecular sequencing (Shaffer et al. (1997), Cervelli et al. 2003; Fujita et al, 2004, Krenz et al 2005, Parham et al. 2006) found the Dermatemydidae and Kinosternidae to be unrelated to the Trionychidae and Carettochelyidae, and reject the monophyletic Trionychoidea sensu Gaffney and Meylan (1988), which becomes a diphyletic taxon in which the Trionychoidea and Kinosternoidea as only very distantly related. A smaller group, Trionychia or Pantrionychia is instead used for the extant Trionychidae and Carettochelyidae and the related fossil forms (the pan- prefix being added by Joyce et al (2004) to include the stem group), these are considered totally unrelated to all other recent Cryptodira (Böhme, 2005, p.410) for which Danilov & Parham, 2006 coined the clade Durocryptodira. Most of these analyses place the Trionychia as the basalmost clade within the cryptodires (Shaffer, 2009). As regards the pattern of chelonian cranial arterial circulation, the Trionychoidea and Kinosternoidea differ from the more widespread Testudinoid condition. This supports a monophyletic Trionychia/Trionychoidae, whereas the more inclusive Trionychoidea taxon sensu Gaffney and Meylan is not supported (Jamniczky & Russell, 2007). In the present work we've followed the molecular phylogenetic position, as this is supported by stratigraphic priority of Trionychoidea, which appear in the fossil record some fifty to a hundred million years before any non-trionychoidan crown cryptodires MAK130302
 |
| Reconstruction of the shell of the basal trionychoidan Basilochelys macrobios from the Latest Jurassic or Earliest Cretaceous of Thailand. Length 90 cm. From Tong et al. 2009 p.162 |
Horizon: latest Jurassic-earliest Cretaceous Phu Kradung Formation of NE Thailand
Trionychidae : (Trionychinae + (Cyclanorbinae + Plastomeninae)) + *
Comments: This large cryptodire (shell length upto 90cm) known from skull, shell and other postcranial elements, would seem to be the most basal member of the Trionychoidea, with a combination of primitive eucryptodire and derived characters. It constitutes a transitional form with many features shared with both Xinjiangchelyidae, Adocidae and Nanhsiungchelyidae. The amphicoelous cervical vertebrae are like those of Xinjiangchelys and Thalassemys, although it differs from the xinjiangchelyids and plesiochelyids by a derived character (posterior portion of the canalis caroticus internus is covered by bone), a Centrocryptodiran synapomorphy. There are also several characters shared with Early Cretaceous Sinemydidae and Macrobaenidae, such as Ordosemys. Several features of the shell are shared with Trionychoidea and Testudinoidea, and the first thoracic vertebra has an anteroventrally facing anterior central articulation, which is a feature of the neck retraction mechanism characteristic of advanced cryptodires. This implies these large turtles were probably capable of retracting the head inside the shell, an advanced feature in cryptrodire evolution (Tong et al. 2009 pp.164-5) Tong et al. 's cladistic analysis resulted in a single tree of 177 steps with a consistency index of 0.47 and retention index of 0.74, suggesting rampant homoplasy in the dataset. The combination of primitive and derived characters in Basilochelys suggests that the Trionychoids may have originated from xinjiangchelyids and their close relatives (Tong et al. 2009 p.168), which supports the molecular and stratigraphic evidence of an early divergence for this group. Cladistic analysis by Syromyatnikova 2012 interprets as a basal Adocusia rather than stem Trionychoidea. In any case, given the ambiguity and homoplasy of the material, any interpretation either way must only tentative. For now we have followed Tong et al. in placing Basilochelys as a stem trionychoidan (Trionychoidea in their paper). MAK130211
Adocusia: the most inclusive clade including Adocus beatus (Leidy, 1865) and Nanhsiungchelys wuchingensis Yeh, 1966
Range: Late Jurassic to Oligocene of Asia, Late Cretaceous to Paleocene of North America.
Phylogeny: Trionychoidea : Basilochelys + ((Peltochelys + Trionychia) + * : Adocidae + Nanhsiungchelyidae + Yehguia)
Characters: Sinuous midline plastral sulcus; Osseous connection between the plastron and carapace (evolved independently within Adocusia, Pankinosternoidea, and Pantestudinoidea, the primitive condition for Cryptodira is ligamentous); sculptured shell.
Comment: stratigraphically by far the oldest trionychoidan group. The phylogeny is still disputed. Hennigian cladistic study (Gaffney & Meylan, 1988; Meylan & Gaffney, 1989)) show the Adocidae as the sister group to the clade (Nanhsiungchelys + (Trionychidae + Carettochelyidae)). Statistical-computational analysis instead reveals a monophyletic clade Adocusia (Adocidae + Nanhsiungchelyidae + Yehguia ) as sister group to Crown Trionychoidea (Danilov & Parham, 2006, 2008; Joyce 2007; Sterli, 2010), although not all agree on the monophyly of the Adocusia (Tong et al., 2009). Primitive features of morphology are characteristic of basal level eucryptodires such as the Xinjiangchelyidae (Syromyatnikova 2012) It may be that this is a paraphyletic assemblage of stem forms rather than a true clade (by analogy with Ceratosauria within theropoda; previously considered to be sister taxon to the Tetanura it is now recognised to be a grade of early theropods), in which case, phylogenetically speaking, Adocusia would equal Trionychoidea, and adocusian synapomorphies are actually shared primitive features modified or lost in more advanced trionychoidans. Alternatively, in contradiction to all current parsimony-blased cladograms, if the Adocusia are not trionychoidans but evolved by convergence, this would explain both why the most morphologically primitive Trionychoidea like Kizylkumemys and perhaps Sandownia, are found in the crown group, as well as tidying up the ridiculous number of ghost lineages. MAK130128
Range: late Jurassic (Oxfordian) of China
Adocusia : Adocidae + Nanhsiungchelyidae + *
Comments: Previously included under Plesiochelys, closer analysis revealed this archaic turtle to be the earliest known trionychoidan by Danilov & Parham, 2006. However Lourenço et al 2012 using molecular dating place the divergence between trionychoidans and non-trionychoidan cryptodires at around 144.0 Mya, or in other words the Jurassic-Cretaceous boundary (Lourenço et al 2012 p.502) . They therefore interpret late Jurassic forms like Yehguia as early crown Cryptodira rather than a trionycha
 |
| Shell of Adocus punctatus, image by Othniel C. Marsh, 1890, public domain. Via Wikipedia |
Range: Late Cretaceous to Paleocene (Cenomonian to Danian) of NAm & Asia
Phylogeny: Adocusia : Nanhsiungchelyidae + Yehguia + * : Adocus + Shachemydinae
Characters: Weakened costal ribheads (Danilov & Parham, 2006)
Comments: A group of large fresh-water aquatic turtles from the Late Cretaceous of Asia and North America. In Asia the Adocidae were more numerous than Nanhsiungchelyidae and often dominated Late Cretaceous complexes turtles. They were mainly confined to the humid subtropical climate conditions on the outskirts of Asia with extensive coastal lowlands (Syromyatnikova 2012). The entire surface of the carapace is finely sculptured with a very regular arrangement of minute tubercles. (Scheyer 2007, p.104)
Originally, through the influential classifications of Lydekker (1889) and Zittel (1889), adocids were placed in the family Dermatemydidae with the extant turtle Dermatemys mawii, based on the absence of mesoplastra, the presence of a complete row of inframarginals, and a well-ossified plastron. However, Meylan and Gaffney (1989) argued on cladistic grounds that these are paraphyletic (shared primitive) features among cryptodires, and that most similarities between adocids and Dermatemys mawii should be interpreted as plesiomorphies or homoplasies. They transferred the adocids to the Trionychoidea, a morphology-only clade that is not supported on molecular grounds. As the phylogenetic relationships of adocids can only be determined using fossil skeletal morphology, Danilov & Parham 2008 reanalysed the morphological dataset of Joyce (2007), using a topological constraint that forced the current molecular consensus and the nonmonophyly of Trionychoidea. Their resulting cladograms placed adocids as the sister group to Trionychia, as basal cryptodirans. (Zelenitsky et al)
|
Adocus Cope, 1868 ((monotypal Subfamily Adocinae Cope, 1870)
Range: Late Cretaceous of Kazakhstan, Central Asia, Late Cretaceous to the Late Paleocene in North America North America, upto Eocene of Asia
Phylogeny: Adocidae : Shachemydinae + *
Description: - Hay (1908) p.237-8,
Comments: Medium to large omnivorous-herbivorous freshwater turtles, in large animals the shell could reach up to 80cm. Aquatic and probably a good swimmer, shell delicately sculptured, flat and long, smoothly contoured, the plastron has large openings to allow free motion of the limbs. May have resembled, but not related to, the Mesoamerican river turtle Dermatemys. Jehle, 2006, with which they were for a long time grouped (e.g. Zittel, Eastman, Woodward, et al(1932), Mlynarski 1976). Known from the Late Cretaceous (Coniacian to Maastrichtian) of North America, extending to the Early Palaeocene (Puercan) (Hell Creek Formation and overlying Tullock Formation) of Montana (Scheyer 2007, p.105) continuing to the Late Paleocene . A number of species are known from the Late Cretaceous (early Cenomanian to early Campanian) of central Asia and Kazakhstan. (Syromyatnikova & Danilov, 2009)
Shachemydinae: Khosatzky, 1977,: Ferganemys, Shachemys
Range: Late Cretaceous of Asia
Phylogeny: Adocidae * : Adocus + *
Characters:. entoplastron is shortened and truncated anteriorly. (Danilov & Parham, 2006), some plastral kinesis
Comments: Shachemys and Ferganemys are both characterized by a short, relatively elongated and streamlined skull, while Adocus it is higher and shorter. The orbits are shifted upward, while for Adocus located on the sides of skull. These differences are likely to be dietary: the presence of relatively high skull suggests greater bite force and power over moving objects for Adocus. In Shachemys skharakternoy a reduction or disappearance of neural plates, also found in some Chelidae ( Chelodina and Emydura ) is associated with a reduction of the space for the passage of the longissimus dorsi muscle (m. longissimus dorsi). This means these animals were less able to lunge with the neck, and, thus, does not allow to make sharp and fast attacks its prey.Feeding more likely involved relatively passive capture and swallowing of prey. Underdeveloped neck muscles are also associated with the presence of the hinge at the front of plastron, providing additional protection for the head with weakly mobile neck. Shachemys is only known from coastal areas (Syromyatnikova 2012)
 |
| Basilemys sp., photo © Royal Tyrrell Museum |
Range: Cenomanian to Maastrichtian of Asia (Mongolia, China, and Japan) and Campanian to Maastrichtian of N Am
Adocusia : Adocidae + Yehguia + * : Kharakhutulia + (Basilemys + Nanhsiungchelys + Zangerlia)
Characters: reduced number of phalanges; thelial process absent (Meylan & Gaffney, 1989)
Comments: A uniquely East Asian - North American Cretaceous group of large testudines (with shells generally around 50 to 90 cm in length (Hirayama et al. 2001) characterised by a combination of aquatic and terrestrial features, although a terrestrial lifestyle seems more likely, making them testudinid precursors (Syromyatnikova 2012). They include the genera Zangerlia, Hanbogdemys, Anomalochelys, Nanhsiungchelys and Basilemys. In contrast to the Adocidae, the Nanhsiungchelyidae preferred inland arid climates, most remains being found in eolian deposits, suggesting they live in hot and dry desert landscapes (Syromyatnikova 2012). The morphology is still poorly known, and the systematic position of a number of forms unclear. This is especially true of Asian species, which are described mainly on the basis of a few fragmented materials (Syromyatnikova 2012) As with the Adocids they were originally included under the Dermatemydidae. (Mlynarski 1976) The external surfaces of the shell bones of Basilemys, Zangerlia and Nanhsiungchelys share a regular pock-marked sculpture (Scheyer 2007, p.105), although sculpturing can be distinguished from those of the Adocidae by the relatively bigger and more irregular pits and grooves (Sukhanov et al 2008 p.602). Cladistic analysis shows the Nanhsiungchelyids to be close to the Adocidae. Some cladograms show an early split between the Asian forms like Nanhsiungchelys and the American Basilemys (Gaffney & Meylan, 1988, Joyce & Norell, 2005 ); others have a number of lineages, with Kharakhutulia as the most basal, , implying that the North American Basilemys is derived from Asian ancestors, although it is not clear which lineage Basilemys is closest to (Sukhanov et al 2008, Syromyatnikova 2012). MAK130222
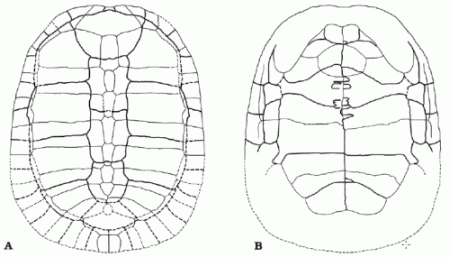 |
| Reconstruction of the shell of the basal Nanhsiungchelyid, Kharakhutulia kalandadzei, in dorsal (A) and ventral (B) views. Length about 25 cm. From Sukhanov et al 2008 p.610 |
Kharakhutulia kalandadzei Sukhanov et al 2008
Horizon: Lower part of the Bainshire Formation, Cenomanian or lower Turonian of Khara Khutul, Mongolia
Nanhsiungchelyidae : (Basilemys + Nanhsiungchelys + Zangerlia) + *
Comments: The most basal Nanhsiungchelyid, smaller than most other members of the family (shell only 23 to 25 cm long), in overall morphology transitional between Adocidae and more advanced members of the Nanhsiungchelyidae (Sukhanov et al 2008 p.610) MAK130205
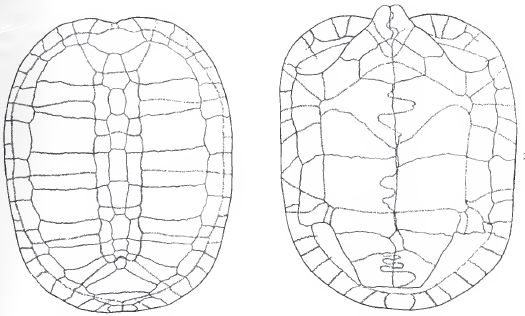 |
| Basilemys sinuosa. Carapace and plastron, length 71.5 cm, breadth 58.5 cm. From Hay (1908) p.229, figs 288 and 289; public domain (copyright expired). |
Range: Late Cretaceous (Campanian to Maastrichtian) of North America
Nanhsiungchelyidae : Kharakhutulia + (Nanhsiungchelys + Zangerlia + * )
Description: Shell highly sculptured. Epiplastra much thickened. Bridge wide. Intergular scutes present. Pectorals greatly expanded at the midline, narrowed laterally. An axillary and an inguinal scute but no other inframarginals - Hay (1908) p.226
Comments: A specialised Laurentian lineage of Nanhsiungchelyids, evolving in isolation from the various Asian forms. The pock-marked sculpturing extends into the interior of the shell openings, with the internal surfaces of bones adjacent to these openings also heavily sculptured. Several morphological features, such as robust elephantine limbs and stubby feet, ossified limb armour, strong epiplastral projections, and triturating surfaces of maxilla, show that Basilemys were the ecological equivalent of terrestrial and herbivorous tortoises (Scheyer 2007, p.105). This distinguishes it from the predominantly freshwater semi-aquatic turtles of the time. However the shell was low and sleek rather than domed as with the testudinidae (Acorn 2007, p.14). Several species are known; Basilemys variolosa is from the Campanian and B. imbricaria, B. nobilis, and B. sinuosa from the Maastrichtian. The latter is known from a nearly complete carapace and plastron holotype from the Maastrichtian of Montana. and also known from North and South Dakota, and Wyoming, had an average carapace of 70 by 56.5 cm, giving an estimated body mass of 38.5 kg Paleobase. The largest specimens of this genus have shells exceeding 1 meter in length. Their large size and terrestrial habitat meant that these was among the few testudine groups that succumbed to the K/T mass extinction (Jehle, 2006).
A purported Asian species, B. orientalis Sukhanov and Narmandakh, 1975, from the Bainshire Formation [upper Turonian or Santonian] of Baishin Tsav, Mongolia (Sukhanov et al 2008), turned out to be unrelated and has been given its own genus, becoming Hanbogdemys orientalis. Hanbogdemys is common in Asia in the range of Cenomanian - Campanian (Syromyatnikova 2012). A single nuchal from Khara Khutul (Cenomanian or lower Turonian) may belong to a distinct but related species (Sukhanov et al 2008 p.612), perhaps an ancestral form. MAK130205
Range: Nanhsiungchelys wuchingensis from the Maastrichtian of China. Related form (Anomalochelys angulata) from Cenomanian of Japan
Nanhsiungchelyidae : Kharakhutulia + (Basilemys + Zangerlia + *)
Comments: A distinctly Asian lineage with a relatively large skull and long tubular snout, Lucas, 2001, pp.182-3 suggests this was likely for rooting out food like the extant tropical Australasian pig-nosed turtle Carettochelys. A more likely explanation is an adaptation to arid climate from the Asian end-Cretaceous. A similar structure is known for the Aldabra giant tortoise Aldabrachelys from the Seychelles. Large nasal cavities which are adapted to the absorption of water through the nostrils which was adapted to living in arid conditions. Syromyatnikova 2012). Anomalochelys angulata Hirayama et al. 2001 from the Cenomanian of Japan (Hirayama et al. 2001) seems to be a closely related form (Sukhanov et al 2008 p.602), indicating that this was a long lived lineage throughout East Asia. MAK130222
 Fossil of Zangerlia neimongoliensis, at the Musee d'Histoire Naturelle, Brussels. Photo by Ghedoghedo Creative Commons Attribution, Wikipedia |
Range: Campanian of Mongolia
Nanhsiungchelyidae : Kharakhutulia + (Basilemys + Nanhsiungchelys + * )
Comments: A primitively or secondarily small genus known from several species (Sukhanov et al 2008). Zangerlia may have fiolled the same ecological role as small testudinids. A number of species are placed here, although this may be a paraphyletic or polyphyletic taxon. MAK130301
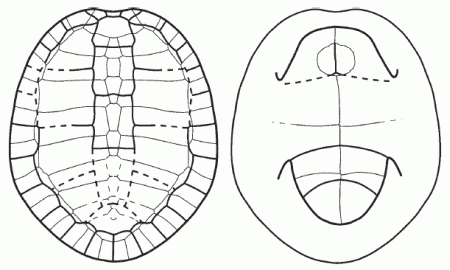
 Peltochelys duchastelli, Early Cretaceous of Beligium. Shell in Dorsal (left) and Ventral (right) view. Drawing from Böhme, 2005, p.413. |
Peltochelys duchastelli Dollo, 1884
Range: Wealden (Barremian/Aptian) of Belgium
Trionychoidea : Basilochelys + (Adocusia + (Trionychia + * ))
Comments: A basal trionychoidan, and the oldest representative of the Trionychoidea outside Asia. The clade Trionychia + Peltochelys is supported by a reduction of the number of peripherals to 10 pairs (Anquetin, 2009 p.204). It shows that this clade had diverged from Adocusian trionychoidans by or around the Barremian age
 |
| Florida Soft-shell Turtle, Apalone ferox, as seen in Everglades National Park, Homestead, Florida, USA.. Photo by Johnskate17, GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
Range: Mid Cretaceous (Aptian) to Recent.
Trionychoidea : Basilochelys + (Adocusia + (Peltochelys + * : Trionychidae + Carettochelyidae ))
Characters: peripherals not sutured to costals; ribheads strongly sutured to vertebral centra; premaxillae fused; radius and ulna in contact adjacent to manus; number of clawed digits three or fewer; coracoid longest of three pectoral processes; carapace not sutured to plastron (Meylan & Gaffney, 1989)
Further description: "Dorsal vertebrae and ribs fused with the dermal plates to form an incompletely ossified carapace ornamented with coarse vermiculating sculpture; no epidermal shields. Neck bending by a sigmoid curve in e vertical plane. Skull with descending parieto-pterygoidal processes. Pterygoids broad throughout, separated from each other, the basisphenoid joining the palatines. Cervical vertebrae without transverse processes. Sacral and caudal ribs generally attached to well-developed transverse processes of the neural arches. Pelvis free from the carapace and plastron. Limbs modified into paddles, the fourth digit with at 1not four phalanges, and only the three inner digits clawed. Marginal hones absent or forming an incomplete series, not connected with the ribs. Nine plastral elements; epiplastra separated from the hyoplastra by the V-shaped entoplastron." - Zittel, Eastman, et al (1902) pp.195-6
Comments: [1] "The group of three-clawed mud-turtles, which appears first in the Upper Cretaceous of the United States, and next in the lowest Tertiary strata of both Europe and North America, exhibits the most generalised structure of all Chelonians. The shell is incompletely ossified, and the plastral elements remain separate throughout life. Vacuities persist in the carapace, and various portions of the skeleton afford evidence of imperfect ossification. There are no free nasals, and no parieto-squamosal arch ; the descending processes of the prefrontal may or may not be connected with the vomer, the epipterygoids are free, and the dentaries distinct. The stapes is entirely surrounded by the quadrate. The pterygoid is broad, without lateral expansions, separating the quadrate and basisphenoid. Only one family is recognised." - Zittel, Eastman, et al (1902) pp.196
[2]. Lourenço et al 2012 using molecular dating place the divergence of crown Trionychoidea into Carettochelyids and Trionychids at 128.7 Mya, or Barremian age of Early Cretaceous.
| The Pig-nosed Turtle Carettochelys insculpta, Dvur Králové Zoo, Czech Republic. Photo by Karelj. Public domain. via Wikimedia |
Range: Mid Cretaceous (Albian) to Recent of Asia
Trionychia : Trionychidae + *
Characters: paired articulation between 8th vertebra and the nuchal bone, limbs terminate in paddles. (Craniata/Anapsida/Anapsida-TAXONOMY.htm">Jack R Holt)
Comments: Medium-sized, freshwater turtles Pignose Turtles. The carapace is leathery; and they have an elongate, pig-like snout. The fossil record of the group is almost exclusively found in the Northern Hemisphere, with fossil forms from North America, Europe, and Asia. Divided into the more basal Anosteirinae with Kizylkumemys Nessov, 1977, Anosteira Leidy, 1871 and Pseudanosteira Clark, 1932, and the larger and more advanced Carettochelyinae which includes Carettochelys Ramsay, 1887, Allaeochelys Noulet, 1867, Burmemys Hutchison et al., 2004 and Chorlakkichelys. The oldest representative of Carettochelyidae is Kizylkumemys schultzi from the middle Cretaceous (Albian and Cenomanian) of central Asia. It also has a number of primitive features. "In 2003, I. G. Danilov and A. O. Averianov (St. Petersburg, Russia) presented new data on the morphology of Kizylkumemys, which appears to be more primitive than Carettochelys with similarities to trionychoid turtles. The new data supports the monophyly of the (crown) clade Trionychia (Carettochelyidae + Trionychidae), although the carotid morphology of Kizylkumemys is closer to the baenid condition (internal carotid canal is floored anteriorly, no foramina basisphenoidales present), which argues for the Trionychia outside of Eucryptodira." (check quote) Fossil Turtle Newsletter Jan 2004 p.7. This supports molecular phylogeny, which posists a very early split between the (pan)Trionychia and all other extant cryptodires. Even older material, mentioned in Hirayama et al. (2000), from the Early Cretaceous (Hauterivian) of Japan, likely represents the phylogenetic stem of Carettochelyidae. During the Eocene, Carettochelyidae had its greatest diversity in Asia, Europe and North America. Today, the only surviving species is Carettochelys insculpta Ramsay, 1887, found in freshwater and brackish water rivers and lagoons in New Guinea and Australia. (Joyce et al 2004, p.1002; Scheyer 2007, pp.106-7) MAK130222.
 |
| Trionyx messelianus from Messel Pit Fossil Site (Middle Eocene of Germany) at the Naturmuseum Senckenberg. T. messelianus can reach a length of about 60 cm and it is the largest species of turtles of Messel Pit Fossil Site. Carapace and plastron are not connected by bony prominences and the plastron show bony plates reduced to small protrusions. According to see Karl (1998) Trionyx messelianus is a synonym of Rafetoides austriacus (Peters, 1858). - Wikimedia. Photo by Hectonichus, GNU Free Documentation/Creative Commons Attribution license, via Wikimedia |
Range: Mid Cretaceous (Aptian) to Recent,
Trionychia : Carettochelyidae + * : Basilochelys + ((Plastomeninae + Cyclanorbinae) + Trionychinae)
Characters:. [1] Fleshy lips over the keratin beak, no articulation between the centra of the last cervical and first thoracic vertebrae, temporal region open, ; webbed feet, claws only on the middle three digits (hence the name - "Trionyx" - Three Clawed). Most of the carapace is lost, replaced by flexible leathery covering instead of scutes; the plastron is also reduced, with the connection through ligaments instead of as bony bridge. The shell is very flattened and the head has elongate rostra (nostrils in a projecting snout)." - Jack R Holt
Further description: "Skull depressed, the small orbits directed more or less upwards and approximated towards the nares; temporal fossae completely open, and squamosal and supra-occipital with very long posterior processes. Plastron totally distinct from the carapace, with large vacuities. Humerus much curved." - Zittel, Eastman, et al (1902) pp.196
Comments: Freshwater riverine (rarely brackish water) mainly carnivores turtles feeding on fish, crustaceans, and insects, fast swimmers. The neck and snout are elongated for prey ambush. They lay 4 to 100 brittle, spherical eggs ref. Trionychidae originated in Asia in the late Early Cretaceous, Aspideretoides allani, Aspideretoides foveatus, Aspideretoides splendidus, and Apalone latus from the Late Cretaceous (middle Campanian) of North America are likely trionychids, thus extending the age of the crown back to the lastest Cretaceous. (Joyce et al 2004, p.1002, Scheyer 2007, p.109). During the Tertiary they spread to all other continents with exception of Antarctica. Most fossil forms placed in the extant genus Trionyx have been described from Eocene and Miocene Europe, although there is the tedemcy now to give them distinct generic names. Axestus and Plastomenus Cope, from the Eocene of Wyoming and New Mexico, are closely related genera. Chitra Gray is known from the Pliocene and Recent (Zittel, Eastman, et al (1902) pp.196).
23 species and 14 extant genera of extant soft-shelled turtles are known from temperate and tropical areas of North America, Eurasia, Africa and New Guinea (Joyce & Lyson 2010 p.241)
 |
| Fossil of Plastomenus sp. in the Field Museum of Natural History, Chicago. Photo by Smokeybjb, GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
Plastomeninae: Hutchemys, Plastomenus, ?Helopanoplia
Range: Late Cretaceous ( Campanian) to Eocene of North America (possibly also Kazakhstan)
Trionychidae : Basilochelys + (Trionychinae + (Cyclanorbinae + *))
Characters:. Well ossified plastron and the central fontanella of the plastron is small or even absent
Comments: Phylogenetic analysis reveals that plastomenids are stem cyclanorbines, either an early branching clade of stem Cyclanorbinae, or a paraphyletic grade that gave rise to modern cyclanorbines. This significantly reduces the previous ghost range (in this case the result of a research bias rather than absense of fossils) of the total group Cyclanorbinae from some 80 to about 30 million years. Although abundant, their fossil record is still too poorly understood. Joyce & Lyson 2010. As the Plastomenidae are now includsed in the total clade Cyclanorbinae, we have downgraded them from family to subfamily ranking (alternatively the Cyclanorbinae can be elevated to family rank).
References: Scheyer 2007, pp.110; Joyce et al 2009, Joyce & Lyson 2010
 |
| Lissemys punctata (Indian flapshell turtle). Illustrations of Indian Zoology - volume 2, Thomas Hardwicke. Public domain. via Wikimedia |
Range: Miocene to Recent
Trionychidae : Basilochelys + (Trionychinae + (Plastomeninae + * ))
Characters:. massively ossified shell and fused hyo- and hypoplast
Comments: Evolved from plastomenine ancestors Joyce & Lyson 2010. Three recent genera - Cyclanorbis, Cycloderma, and Lissemys - with two species each. Lissemys still retains small bone elements in the periphery of the soft dermal part of the shell, and is considered the most basal extant representative of the Trionychidae. The shells are also more extensively ossified than trionychines, a set of traits that is generally considered to be morphologically less specialised (and hence closer to the ancestral condition). Joyce & Lyson 2010 p.241The subfamily is known in the fossil record from the Miocene of Saudi Arabia and the Pliocene of Africa and India. (Scheyer 2007, pp.110)
 |
| Eastern Spiny Softshell Turtle (Apalone spinefera). Photo by Kim Pardi, Creative Commons Attribution license, via Wikipedia |
Trionychinae: Extant genera Amyda, Apalone, Aspiderestes, Chitra, Dogania, Nilssonia, Palea, Pelochlys, Pelodiscus, Rafetus, Trionyx.
Range: Mid Cretaceous (Aptian) to Recent
Trionychidae : Basilochelys + (Cyclanorbinae + Plastomeninae) + * )
Characters:. plastral bones not firmly fused but loosely bonded by connective dermal tissue
Comments: Medium-sized, freshwater turtles, known from the Mid Cretaceous of North America onwards (Scheyer 2007, pp.110-111). In contrast to cyclanorbines, trionychines exhibit a number of traits that are typically interpreted as derived, including "a reduction in the ossified carapacial disk to the point that the pelvis is not covered by bone, the resulting development of an extensive leathery carapacial perimeter with the formation of extensive free rib ends, and the reduction in the plastral callosities, particularly those of the entoplastron and epiplastra" Joyce & Lyson 2010 p.241
Page last modified MAK130301.
All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners

{kind=link}
{kind=link}