| Chelonii | ||
| The Vertebrates | Chelonioidea |
| Vertebrates Home | Vertebrate | Vertebrate |
Taxa on This Page
The Chelonioidea or sea turtles are among the most distinctive of reptiles, and have been characteristic of marine ecosystems since the late Cretaceous. They are easily distinguished from other testudines by their flipper-shaped limbs. with the fore-pair the larger and low streamlined shell. As with some freshwater aquatic forms like the Trionychoidae there is the tendency to reduce the bony shell, so that in the leatherback turtle this has becomes little more than numerous bony platelets situated in thick, leathery skin. Unlike other cryptodires, they cannot retract their head into the shell. As with the Mesozoic ichthyosaurs and plesiosaurs, the marine turtles have a worldwide pelagic distribution.
Despite the strong similarity with the Cretaceous Protostegids, the Chelonioidea are only very distantly related; both groups independently evolved paddles, a reduced, streamlined shell, and large size from different semi-aquatic terrapin-like ancestors. The dramatic evolutionary radiation of the chelonioids during the Late Cretaceous (Late Campanian age) may be due to the availability of ecological opportunities vacated by protostegids, which had became extinct by this time (Parham & Pyenson 2010 p.242). During the latest Cretaceous through the Cenozoic they diversified into a range of ecological roles. Now after 75 million years of highly succesful evolution, these large, charismatic, and highly specialised reptiles, like every other species of large wild animal, are themselves facing extinction due to human predation, poaching, and ecological enroachment MAK130224
|
|
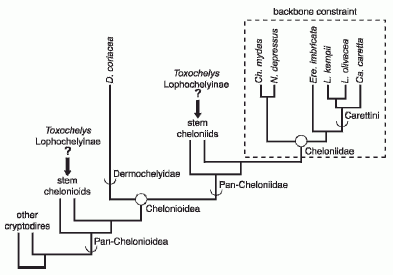
| Phylogenenetic taxonomy (Joyce et al 2004 and later) and cheloniid molecular sequenced (Naro-Maciel et al 2008 and later) constraint tree. Full circles represent node-based definitions whereas semi-circles represent stem-based definitions. showing the uncertain position of the fossil taxa Toxochelys and the Lophochelyinae as either stem cheloniids or stem chelonioids; the same ambiguity applies to other fossil taxa e.g. Mesodermochelys, Allopleuroninae. From Parham & Pyenson 2010 fig.1 |
Panchelonioidea (= Panchelonioidea of Joyce et al 2004, = Superfamily Chelonioidea) : Sea Turtles and their fossil relatives
Range: Cretaceous to Recent
Phylogeny: Chelonioidea-Chelydridae-Kinosternoidea clade : (Chelydridae + Emarginachelys + Kinosternoidea) + * : Chitracephalus + Chelonioidea
Characters: "xiphiplastral fontanelle, which involves the reduction of the xiphiplastra in narrow struts that frame it" - Pérez-García 2012 p.577,
Comments: The name Panchelonioidea is used by Joyce et al 2004 to refers to the clade that includes both extant Chelonioidea and a number of extinct and ancestral lineages. It basically means Chelonioidea as usually defined. The Pan- prefix as a signifier of a phylogenetic stem group and crown taxa. As yet it has not caught on outside testudine studies, and we have retained the Linnaean superfamily for the sake of consistency. MAK130301
 |
| Chitracephalus dumonii, from sketch in Pérez-García & Murelaga (2012). Note the very long neck and almost circular shell |
Chitracephalus dumonii Dollo, 1885
Synonym: Salasemys pulcherrima
Range: Hauterivian/Barremian to Barremian/Aptian of Belgium and Spain.
Phylogeny: Panchelonioidea : Chelonioidea + *
Description: "Cranium much elongated and depressed, with an extremely short facial region, and temporal fossae not roofed. Cervical vertebrae without transverse processes. Pleurals narrowed at their outer ends, with vacuities within the well-developed border of the peripherals. Plastral elements similar to those of Chelone. Digits clawed, not greatly elongated." - Zittel, Eastman, et al (1902) pp.198
Comments: Originally of uncertain placement (classification Testudinata incertae sedis) cladistic analysis of this long necked semi-aquatic turtle by Pérez-García 2012, shows that Chitracephalus is actually a basal Chelonioidean. Together with Hoyasemys, Chitracephalus forms part of an Early Cretaceous European clade that includes macrobaenid, sinemydid, and panchelonioidean turtles. MAK130306
 |
| Carapace terminology, showing Chelonia. The shape, arrangement, and type of scutes and its distinctive jargon is essential for understanding testudine paleontology and phylogeny (see also our Primer on Turtle Shell Anatomy. From G.A. Boulenger, Fauna of British India. Amphibia and Reptilia. 1890, public domain, via Wikipedia |
Chelonioidea : Sea Turtles and their fossil relatives
Range: Cretaceous to Recent
Phylogeny: Panchelonioidea : Chitracephalus + * : (Mesodermochelys + ?Allopleuroninae) + Toxochelyidae + Crown Group Chelonioidea)))
Characters: "raised pedestal on the visceral surface of the nuchal for the articulation with the neural spine of the eighth cervical vertebra present; reduction of costal ossifications, central plastral fontanelle retained in adult individuals; epiplastra elongate in shape; and paddles present" - Anquetin, 2009 p.202. "well developed costal fontanelles" Pérez-García 2012 p.577, Anquetin, 2009 p.202
Comments: The precise phylogenetic position of Cretaceous taxa like Toxochelys and Mesodermochelys is uncertain, but the general consensus now is that they represent stem taxa (with Mesodermochelys and Toxochelys forming a polytomy with Crown Group Chelonioidea (Anquetin, 2009 p.202), or Mesodermochelys may be more basal than Toxochelys (Pérez-García 2012) rather than early representatives of the Cheloniidae and Dermochelyidae. MAK130301
Links: Wikipedia; Oceans of Kansas (late Cretaceous forms);
 |
| Toxochelys?, Smithsonian National Museum of Natural History: Hall of Fossils Photo by Ryan Somma, via Wikipedia, with caption, Creative Commons Attribution-Share Alike 2.0 |
Toxochelyidae Baur (1896) (ref)
Range: Late Cretaceous (Coniacian to Earliest Maastrichtia) of North America
Phylogeny: Chelonioidea : (Mesodermochelys + ?Allopleuroninae) + Crown Group Chelonioidea + * : Toxochelys + "Thinochelyinae"
Description: "Skull somewhat depressed. (flat) Temporal region extensively roofed. Quadrate notched for the columella. Choanae situated well forward; not underfloored by the palatines. Palatines extending forward to the vomers and forming the outer boundaries of the choanae. Hay (1908) p.163 Anterior vomer area wide and smooth. Carapace circular, rarely cordiform.(Zangerl, 1953b p.173) with eleven pairs of peripherals, in addition to the nuchal and pygal. Neuralia flat (i.e. no keel along the bones of the midline of he carapace). Nuchal not furnished with costiform processes. Epidermal shields present. Plastron loosely articulated with the carapace, not extending forward to the third peripheral and backward hardly to the eighth. Fore foot with at least 2 claws; the phalanges furnished with condyles; the limb as a whole resembling that of the Trionychidae."
Comments: A group of late Cretaceous near shore marine turtles that show a number of primitive features that distinguish them from the Cheloniidae. Although some cladistic analyses show Toxochelys (and by implication related Toxochelyidae) to be stem-cheloniids (e.g. Hirayama 1998), the consensus now is that they represent a lineage of stem chelonoidea, prior to the cheloniid-dermochelyd divide (e.g. Gaffney & Meylan, 1988, Kear & Lee 2006, Anquetin, 2009, Pérez-García 2012), They seem to have been geographically limited to the American Cretaceous inland sea. As well as Toxochelys, other genera like Porthochelys, and Thinochelys have also been placed here. Toxochelys is known from several species. These turtles were probably not particularly good swimmers; unlike the Cheloniidae the shell is not excavated above the forelimbs (Zittel, Eastman, Woodward, et al (1932), p.316), so there is more limited range of motion. Their eyes face to the top, not outward and slightly forward as in modern sea turtles. These animals may have been bottom dwellers in very shallow sea, perhaps somewhat comparable in habits to the soft-shelled and snapping turtles of freshwater streams and ponds. (Jehle, 2006). Falling sea levels during the early Maastrichtian may have been an environmental factor contributing to their demise. MAK130310
Range: Late Cretaceous (Coniacian to Earliest Maastrichtia) of North America
Phylogeny: Toxochelyidae :
Description: Like Cheloniidae except no secondary palate, fore-foot with at least two claws, phalanges with condoyles (Zittel, Eastman, Woodward, et al (1932), p.316); "Skull longer than broad. No nasal bones. Carapace with large lateral fontanels. None of the peripherals in contact with the disk of the carapace. Midline with prominent carina rising at intervals into tubercles, some of which are distinct bones. Tail with a series ot compressed tubercles above. Plastron with median and lateral fontanels."- (Hay (1908) p.163)
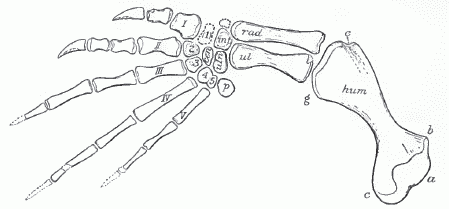 |
| Toxochelys latiremis. Front limb. x 0.28 life-size. Specimen in Yale Univ. collection. a, head of humerus; b, radial process; c, ulnar process; cen, centrale; e, ectepicondylar groove; g, entocondyle; hum, humerus; intj, intermedium; pj pisiforme; rad, radius; ul, ulna; uln, ulnare; I, II, etc., metacarpals; 1, 2, 3, etc., distal carpal bones. Figure by Wieland. From Hay (1908) p.166, fig 200; public domain (copyright expired). The limb is only slightly paddle-like, unlike more specialised marine turtles like Archelon or Carretta |
Comments: This genus gives its name to the family Toxochelyidae. The type species Toxochelys latiremis Cope 1873 is the most common fossil turtle in the Smoky Hill Chalk, Niobrara Formation, western Kansas, of late Cretaceous Santonian age (at the time a shallow inland sea). Although some specimens may only be 15 cm in length (Mlynarski 1976, p.41), these obviously reprent juveniles, adults having a carapace of around a meter in length (Hay (1908) p.169; Oceans of Kansas ref). It was contemporary with Ctenochelys, which was closer to modern turtles.
The Early Campanian T. moorevillensis Zangerl, 1953 from the Mooreville Chalk, Alabama was convergent on the modern Green Sea Turtle Chelonia and on a number of distinct fossil forms in having thin dentaries with large symphyseal ridges (Zangerl, 1953b p.176) ; it was probably the first of the herbivorous chelonoids, associated with the evolution of sea grasses (Parham & Pyenson 2010). The stratigraphically oldest species of Toxochelys is T. latiremis from the Niobrara, is also the most highly specialized. The less specialised Early Campanian T. moorevillensis, T. barberi Schmidt; from the late Campanian Marlbrook Marl, Arkansas, and T. atlantica Zangerl, 1953 from the Upper Greensand, New Jersey.represent a series that occur along the open waters of the Gulf and Atlantic coasts, whereas T. latiremis lived in a semi-isolated inland basin, and hence may have underwent specialization earlier and at a faster rate than its relatives along the Gulf coast (Zangerl, 1953 p.270). T. browni Hay is a very large species from the Early to Mid Campanian Pierre Shale, South Dakota and Wyoming is much larger than T. latiremis. here is some disagreement regarding synonymy, the Paleobiology Database includes every species but under Toxochelys latiremis) Toxochelys weeksi Collins; Ripley Formation, Tennessee.; from the earliest Maastrichtian, is the last known member of the genus (Karl & Lindow, 2009).
Link: Oceans of Kansas.
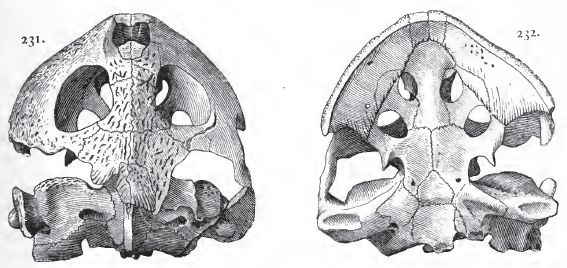 |
| Porthochelys laticeps. Skull of type, half life size. Niobrara formation (Santonian), Kansas. From Hay (1908) p.181, figs 231 and 232; public domain (copyright expired). |
Porthochelys laticeps Williston
Range: Santonian to Early Campanian of North America
Phylogeny: Toxochelyidae : Toxochelys + *
Description: "Skull as broad as long (right). Nasal bones present. Lateral fontanels of the carapace obsolete. All of the peripherals articulating closely with the costals. No carina along the middle of the back and no tubercles. Plastron with small median and lateral fontanels." - Hay (1908) p.180
Comments: Like Toxochelys, Porthochelys seems to be a near coastal species, in contrast to the protostegoids (Hay (1908) p.38). The skull of Porthochelys is much more robust and much wider than that of Toxochelys. The strength of construction and width are correlated with the greater masticatory surface areas on upper and lower jaws (Zangerl, 1953 p.173). The different-shaped head implies different feeding strategies and lifestyle, hence the co-existence of these two similar species. As is so often with fossil forms, it is likely that the many named species are junior synonyms of the type, with differences due to intra-specific variation and post-mortem and geological distortion.. This was a reasonably large animal, the nearly circular carapace of the type specimen being 78.5 cm wide and 73 cm long Hay (1908) p.182
The Informal name "Thinochelyinae" (Mikko's phylogeny) might be used for several species, especially Porthochelys laticeps Williston, 1901, Thinochelys lapisossea Zangerl, 1953, assuming they are more closely related to each other than Toxochelys. MAK130310
Mesodermochelys undulatus Hirayama & Chitoku, 1996
Range: Santonian to Early Maastrichtian of Japan
Phylogeny: Chelonioidea : Toxochelyidae + Crown Group Chelonioidea + (?Allopleuroninae + * )
Comments: Mesodermochelys undulatus is usually considered a Cretaceous relative or ancestor of Dermochelys coriacea (e.g., Hirayama & Chitoku, 1996; Hirayama 1998, Kear & Lee 2006). Cladistic analysis by Joyce, 2007 found only very weak support for this, in the form of partial loss of carapacial scales. Anquetin, 2009 p.202 analysis recovered Toxochelys latiremis, Mesodermochelys undulatus, and Crown Chelonioidea as an unresolved polytomy at the base of panchelonioids, while Pérez-García 2012, incorporating Chitracephalus dumonii in a modified data set of Joyce (2007) found Mesodermochelys to be even more basal than Toxochelys.
Mesodermochelys firest appears in the early part of the Late Cretaceous. An unnamed primitive species with a carapace of less than a meter long was the most dominant sea turtle known from the Santonian of Hokkaido Prefecture, Japan. The larger Mesodermochelys undulatus from the Campanian to Early Maastrichtian of Japan (Hirayama 2012). As with more specialised sea turtles, Mesodermochelys had elongated front flippers. This species is particularly abundant in the Maastrichtian Hakobuchi Group of Hokkaido Prefecture, although it is also known from the Late Campanian to Early Maastrichtian Izumi Group of Hyogo and Kagawa Prefectures, western Japan. This was a large animal, with a carapace reaching 1.3 to 1.5 metres in length. An isolated humerus is about 50 cm long, suggesting an individual with almost 2 m long carapace. (Hirayama 2012) Only the neural or spinal scutes, or individual plates, of the carapace are well grooved. (Wikipedia) MAK130301
Range: Maastrichtian of Europe.
Phylogeny: Chelonioidea : Toxochelyidae + Crown Group Chelonioidea + (Mesodermochelys + * : Allopleuron + Protosphargis)
Comments: The subfamily Allopleuroninae is sometimes used to include a number of Late Cretaceous to Early Cenozoic genera (Allopleuron, Eosphargis, Glyptochelone and Protosphargis) (Bishop 2007) considered early Dermochelyidae. Of these, Eosphargis can confidently be placed in the Dermochelyidae, but the others are of iuncertain phylogenetic position. They may be specialised stem Cheloniids convergent with Dermochelyids, or perhaps equally or more likely a distinct line or lines of stem Chelonoideans. Parham & Pyenson 2010 informally place Allopleuron hoffmanni near Mesodermochelys undulatus. As well as being near contemporaries, both have a reduced, Dermochelyid-like shell and thin dentaries with large symphyseal ridges which imply a possibly herbivorous diet (Parham & Pyenson 2010), although of course these could easily have simply been convergences. These turtles replaced the Toxochelys moorevillensis, which disappeared around the end of the end of the Campanian. MAK130310
 |
| Allopleuron hoffmanni, at Natural History Museum of Maastricht, photo by Ghedoghedo, Creative Commons Attribution license, viavia Wikimedia |
Allopleuron hoffmanni (Gray, 1831)
Range: Late Maastrichtian of Netherlands
Phylogeny: Allopleuroninae : Protosphargis + *
Description: Carapace long and narrow, nuchal deeply emarginate, neurals short and wide with a long keel; marginals long and slender. Zittel, Eastman, et al (1932) p.198
Comments: A fairly large, highly specialised sea turtle, with a plastron length of about 1.2 meters (Mlynarski 1976 p.51). The carapace is greatly reduced, making it unlikely to be an ancestor of Cenozoic cheloniidae. - Hay (1908) p.209. May or may not be related to either or both Protosphargis and Mesodermochelys. Included by Gaffney and Meylan, 1988 in the Euchelonioidea MAK130311
 |
| Protosphargis veronensis Capellini , 1884, from the Latest Cretaceous (Maastrichtian) of Northern Italy. Artwork by Dmitry Bogdanov, GNU Free Documentation/Creative Commons Attribution license, via Wikimedia |
Protosphargis veronensis Capellini , 1884
Range: Maastrichtian of Italy
Phylogeny: Allopleuroninae : Allopleuron + *
Description: Body skeleton similar to that of Protostega, but bones of plastron more slender, and median fontanelle larger. Zittel, Eastman, et al (1902) pp.198
Comments: This animal was probably very similar to today's leatherback turtle ( Dermochelys coriacea), and it is thought that Protosphargis' shell, which could exceed two meters in length, may have been similarily equipped with longitudinal ridges along the back, which help these animals to move quickly in the water. The reduction of the shell is greater than that of the Protostegidae, and more reminiscent of Dermochelys. The bones of the limbs also show a remarkable specialization to marine life. Either a primitive dermochelyid or an unrelated but convergent form. MAK130316
Links: Wikipedia (Italian) Wikipedia (Dutch)
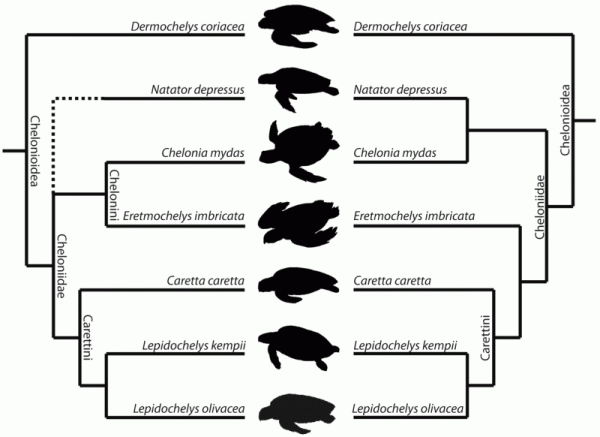 |
| Alternative relationships of chelonioid species according to (left) the morphology-based phylogeny of Gaffney and Meylan 1988 (Natator depressus was not included in that analysis) and (right) the molecular-based phylogeny of Thomson and Shaffer 2010. Names of higher taxa follow Gaffney and Meylan 1988. Silhouettes not to scale. Diagram from Jones et al, 2012 fig 1; Creative Commons Attribution. |
Range: Fr the Late Cretaceous
Phylogeny: Chelonioidea : Toxochelyidae + (Mesodermochelys + ?Allopleuroninae) + * : Pancheloniidae + Dermochelyidae))
Characters: "nasal absent; parietal-squamosal contact present, temporal emargination poorly developed; foramen palatinum posterius absent; and rostrum basisphenoidale rod-like, thick and rounded" - Anquetin, 2009 p.202.
Comments: Gaffney and Meylan, 1988 erected the clade Euchelonioidea for the Cheloniidae, and a clade uniting Notochelone, Allopleuron, Desmatochelys, and the Dermochelyoidae (Dermochelyidae and Protostegidae). The tendency now (according to statistical-computational cladistics) is towards interpreting these Dermochelyid-like taxa as the result of evolutionary convergence. The name Euchelonioidea can be still be used in a more limited context as a synonym to or alternative for the confusing (because the ambiguity in regards to the Linnaean-based taxonomy) phylogenetic use of "Chelonoidea" and the the clumsy "Crown Chelonoidea" MAK130311
Extant Chelonioidea include two families and eight species of extant sea turtles, the monospecific Dermochelyidae (Dermochelys coriacea) and the Cheloniidae. Both lineages are represented by a number of fossil forms from the late Cretaceous and Cenozoic. They are pelagic animals with a worldwide distribution. Diet varies according to species, the giant leatherback turtle (Dermochelys) feeds mainly on jellyfish, ridleys and loggerheads (Caretta and Lepidochelys) on crabs and other benthic invertebrates, and green turtle (Chelonia mydas), the only herbivorous species, adults feeding on kelp [Thalassia testudinium]
Although they are widely protected, every living species is endangered by human activities, whether poaching, oceanic pollution, being caught in nets, injury through boat propellers, or loss of nesting sites through urbanization of coasts and their encasement in seawalls and revetments, and increasingly efficient fisheries.
Links: Geologic History of Sea Turtles, Wikipedia
Pancheloniidae Joyce, Parham & Gauthier, 2004
Range: Fr the Late Cretaceous, pan-oceanic
Phylogeny: Crown Group Chelonioidea : Dermochelyidae + * : Mexichelys + (Lophochelyinae + Cheloniidae)
Characters: (from Gaffney & Meylan 1988): Secondary palate formed by broad maxilla with contributions from palatine, choanal passages separated by ventral plate of vomer forming prominent part of palate; foramen caroticum laterale and canalis caroticus lateralis much larger in diameter than foramen anterius canalis carotici interni and medial branch of canalis caroticus internus. - via Taxon Diversity - CKT
Comments: As with Panchelonioidea, the name Pancheloniidae is used by Joyce et al 2004 to refer to the stem-group that includes crown Cheloniidae and a number of extinct and ancestral lineages (although technically speaking in cladistics you are not supposed to say "ancestors" because it cannot be proved that one species is an ancestor, we are using the term in the colloquial sense). Alternatives are "Chelonioinea" and Cheloniidae Gray, 1825 sensu Hirayama, 1997. We have retained the original Cheloniidae to refer to both more advanced stem and also crown taxa. Cheloniidae in this definition is represented by such taxa as Puppigerus and Euclastes (Joyce et al 2004 p.1001). The boundaries between (Pan)Cheloniidae and more primitive or ancestral stem (Pan)Chelonoidea are blurred. On the one hand, Lophochelyines and Euclastes (as "Osteopygis") have been included in the Toxochelyidae (Carroll, 1988, p.616); on the other, Toxochelys and Ctenochelys from the Late Cretaceous of North America have also been included in the Cheloniidae (Hirayama, 1998). As several recent cladistic analyses shows Toxochelys to be more distantly related (Joyce, 2007, Anquetin, 2009, Parham & Pyenson 2010 fig.3, Pérez-García 2012) , it is here considered basal Chelonioidea. MAK130310
Mexichelys coahuilaensis (Brinkman, Aquillon- Martinez, de Leon Dávila, Jamniczky, Eberth, and Colbert, 2009)
Range: Late Campanian of Mexico.
Phylogeny: Pancheloniidae : (Lophochelyinae + Cheloniidae) + *
Comments: Previously included under Euclastes, this is a much more primitive animal, either a stem chelonioid or a very basal cheloniid, and only distantly reelated to Cenozoic cheloniids . It is distinguished from both both stem-chelonioids such as Toxochelys and stem-cheloniids like Ctenochelys (Lophochelyinae) by an extensive secondary palate, an advanced feature that allows simultanous breathing and feeding. Mexichelys fed on hard-shelled prey (durophagy). These animals were among the first that replaced the protostegids during the Campanian, but by the Maastrichtian they had in turn been replaced by convergently durophagous stem-cheloniid lineage that continued into the middle Miocene. (Parham & Pyenson 2010)
Range: Coniacian to Mid Maastrichtian of North America.
Phylogeny: Pancheloniidae : Mexichelys + (Cheloniidae + * )
Comments: The Lophochelyinae are a family of Late Cretaceous primitive sea turtles that represented a distinct and apparently geographically limited lineage. They range in size from the small Lophochelys, with a plastron length of about 20 cm, to the large Ctenochelys, with an equivalent length of aboout a meter, and the very large Prionochelys, witha shell of about 1.5 meters (Mlynarski 1976 pp.43-4). The fossil remains of these animals have been found in Alabama, Arkansas and New Jersey. They have been various placed in the Toxochelyidae (Zangerl, 1953b, Mlynarski 1976, Kear & Lee 2006), the stem Chelonoidea (Gaffney and Meylan, 1988), and the Pancheloniidae (Parham & Pyenson 2010). We have tentatively followed the latter's phylogeny but it should remain provisional at best. MAK130316
Range: Fr the Latest Cretaceous, pan-oceanic
Phylogeny: Pancheloniidae : Mexichelys + (Lophochelyinae + * : Euclastes + Gigantatypus + Argillochelys + Eochelone + Itilochelys + (Pacifichelys + (Puppigerus + Cheloniinae))
Characters:
Comments: Here used rather informerly to include Euclastes grade sea turtlles and above, which heps to reduce the long phylogenies listed above. There probably isn't a clear distinction between Cheloniidae and Pancheloniidae, unless it's an apomorphic one (defined by specific characteristics not found in Lophochelyines etc) MAK130311
 |
| Lytoloma elegans from the Latest Cretaceous of Morocco. Photo copyright Roger Perkins |
Euclastes Cope, 1867 and related forms
Synonyms: Rhetechelys, Osteopygoides, Osteopygis (crania only)
Range: Latest Cretaceous to Eocene of North America. Lytoloma Cope, 1869 is a closely related genus, it is known from the Latest Cretaceous of England, as well as Morrocco and possibly the Eocene of Tunisia.
Phylogeny: Cheloniidae : Gigantatypus + Argillochelys + Eochelone + Itilochelys + (Pacifichelys + (Puppigerus + Cheloniinae)) + *
 |
| "Osteopygis" (= Euclastes) life reconstruction. Copyright John Maisano |
Characters: Skull resembling that of Caretta. Carapace rounded posteriorly, vacuities of shell smaller than in Carettas; epiplastrals narrow; exposed portion of entoplastron very short, xiphiplastrals uniting extensively in the median line. Zittel, Eastman, et al (1902) pp.198. Broad, flat extensive secondary palate; wide, dentary bone with an elongated symphysis and flat triturating surface; low tomial ridge on the beak, and primitive humerus (Lynch and Parham 2003
Comments: Head broad and flattened, with eyes far forward, like the living loggerhead turtle (Zittel, Eastman, Woodward, et al(1932), p.315). Unlike the contemporary Toxochelys and Eochelone, Euclastes had a secondary palate, although not as developed as in Ctenochelys. The widened palate and dentaries jaws were suitable for crushing hard-shelled organisms, (Lynch and Parham 2003, wikipedia), they represent one of a number of distinct ecologically and morphologically convergent hyper-durophagous cheloniids, the Maastrichtian Euclastes wielandi; (Hay, 1908) replacing the earlier but similar Late Campanian Mexichelys (Parham & Pyenson 2010). The carapace is rounded at the rear, vacuities (gaps in the shell, a common marine turtle feature) not as large as in modern forms (Zittel, Eastman, Woodward, et al(1932), p.315). Lytoloma elegans from the Late cretaceous/Early Paleocene phosphate bed deposits near Khouribga, Morocco is a closely related form. These near-shore and marine turtles inhabited the opening Atlantic coastline of West and North Africa and Western Europe. The well-known Osteopygis is a chimera based on the head of Euclastes and the post-crania of macrobaenids. Durophagous (feeding on hard-shelled invertebrates) 'Osteopygis'-like turtles are known from many localities including South America and Africa (Parham 2005 p. 75).
The genus is something of a wastebasket taxon. Lynch and Parham 2003 attempted to phylogenetically define Euclastes as those taxa that share a more recent common ancestor with the latest Creteacous type species Euclastes platyops Cope 1867 than to extant sea turtles or well-known Paleogene taxa, but the only specimen of E. platyops is too incomplete for phylogenetic analysis (Parham & Pyenson 2010). Parham & Pyenson 2010 assign only three species to the genus, E.. platyops, E. acutirostris Jalil, de Lapparent de Broin, Bardet, Vacant,Bouya, Amaghzaz, and Meslouh, 2009 and E. wielandi, as, unlike other durophagous stem cheloniids, all three show dorsally facing orbits, implying perhaps a specialised, shallow water lifestyle. Other species are transferred to distinct genera, including the new genera Mexichelys and Pacifichelys (Parham & Pyenson 2010), The Mid Eocene Glossochelys planimentum is a specialised durophagous form with a high vaulted skull (Parham & Pyenson 2010 p.235) that may or may not be related, All these stem cheloniids are very similar, and in the past the various genera have been synonytmised (Zittel, Eastman, et al (1932)) MAK130310
 |
| Gigantatypus, at Eternal River Museum, photo by Ghedoghedo, Creative Commons Attribution license, viavia Wikimedia |
Gigantatypus salahi Kaddumi, 2006
Range: late Maastrichtian of Jordan
Phylogeny: Cheloniidae : : Euclastes + Argillochelys + Eochelone + Itilochelys + (Pacifichelys + (Puppigerus + Cheloniinae)) + *
Comments: Known only from an enormous humerus, this gigantic sea turtle lived in the southern regions of the Tethys Ocean about 100-120 kilometres off the north eastern margins of Cretaceous Africa. Estimated total length is over 3.5 metres in length, equivalent to that of Archelon Wieland, 1896. (Parham & Pyenson 2010). We have tentatively followed the latter's phylogeny but it should remain provisional at best. MAK130316
Eochelone brabantica Dollo 1903
Range: Middle Eocene of Belgium.
Phylogeny: Cheloniidae : Euclastes + Gigantatypus + Argillochelys + Itilochelys + (Pacifichelys + (Puppigerus + Cheloniinae)) + *
Comments: This genus lends its name to the paraphyletic subfamily Eocheloniinae, which includes a number of stem cheloniids. The skull of Eochelone is broad and flattened, the eyes are to the side rather than to the front as in more advanced forms. The carapace is elongated and does not have a keel. (Zittel, Eastman, et al (1932)). This was a small sea turtle, with a plastron length of about 60 cm (Mlynarski 1976 p.46)
Itilochelys rasstrigin Danilov, Averianov & Yarkov , 2010
Horizon: Beryozovaya beds (Danian) of Rasstrigin 2 locality, Dubovsky, Volgograd Oblast, Russia.
Phylogeny: Cheloniidae : Euclastes + Gigantatypus + Argillochelys + Eochelone + (Pacifichelys + (Puppigerus + Cheloniinae)) + *
Comments: From Wikipedia: Itilochelys rasstrigin is known only from three fossils, the holotype, specimen number "ZIN PH 1/118" and two referred specimens, numbers "ZIN PH 2/118" and "ZIN PH 3/118" . The type specimen is composed of a grouping of cervical vertebrae I-III, partial skull, and disarticulated lower jaw. Specimen ZIN PH 2/118 is an isolated section of dentary while ZIN PH 3/118 is a right humerus. The fossils were all recovered by A. A. Yarkov from fossiliferous outcrops of the Beryozovaya beds exposed at the Rasstrigin 2 locality.[1] As of 2011, the type and referred specimens are preserved in the Paleoherptological collections housed at the Zoological Institute of the Russian Academy of Sciences, in Saint Petersburg, Russia. Itilochelys was first studied by I. G. Danilov and A. O. Averianov of the Zoological Institute along with A. A. Yarkov of the Natural History, Humanitarian Institute in Volgograd. Their 2010 type description of the genus and species was published in the journal Proceedings of the Zoological Institute RSA. The generic name was coined by Danilov, Averianov, and Yarkov as a combination of the medieval Arabian/Persian name of the Volga River, Itil, along with the Greek "chelys", meaning turtle. The specific epithet rasstrigin is in reference to the type locality of the specimen.
Itilochelys rasstrigin is one of three fossil cheloniid sea turtle taxa which have been recovered from the Volgograd region and the most complete specimen recovered so far. The other two taxa are represented by a specimen of the genus Euclastes known from the Karpovka site and a specimen related to the genus Tasbacka from the Malaya Ivanovka site.
The placement of Itilochelys rasstrigin in the family Cheloniidae is supported by the contact of the squamosal and parietal bones, an emarginated v-shaped crest on the underside of the basisphenoid, and features of the nasal bone structure. Placement of the species as a stem member in the family is based on the structure of the humerus which shows a morphology transitioning towards the derived pelagic structuring of modern cheloniids. (Danilov et al 2010)
 Shell of Argillochelys subcristata and skull of Puppigerus crassicostata, at the Museum für Naturkunde, Berlin. Photo by Ghedoghedo Creative Commons Attribution, Wikipedia |
Argillochelys Lydekker in 1889.
Range: Late Paleocene to Early Eocene of England (A. cuneiceps (Owen, 1849); and A. antiqua); Middle Eocene of Morocco (A. africana (Tong & Hirayama, 2008]).
Phylogeny: Cheloniidae : Euclastes + Gigantatypus + Eochelone + Itilochelys + (Pacifichelys + (Puppigerus + Cheloniinae)) + *
Description: "skull short and wide, shell and bones of the pectoral arch similar to those of Caretta, except that the carapace has but four costal shields, and the xiphiplastrals unite extensively along the median line" - Zittel, Eastman, et al (1902) pp.198
Comments: A small sea turtle, with a plastron length of about 20 cm (Mlynarski 1976 p.48) (unless these fossil forms are huveniles). Argillochelys is the third (after Toxochelys and Allopleuron-Mesodermochelys) of four ecologically convergent specialised lineages with thin dentaries with significant symphyseal ridges imply shearing rather than crushing; possibly feeding on plants like sea grasses. (Parham & Pyenson 2010)
Pacifichelys Parham & Pyenson 2010
Range: Miocene of North America.
Phylogeny: Cheloniidae : Euclastes + Gigantatypus + Argillochelys + Eochelone + Itilochelys + ((Puppigerus + Cheloniinae) + * )
Comments: Originally described as Euclastes hutchisoni is the youngest member of this lineage and is known from the middle Miocene Temblor Formation of California. (Lynch and Parham 2003, Brinkman 2009 p.39). Later phylogenetic analysis by (Parham & Pyenson 2010 shows Pacifichelys to be a more advanced form, closer to Puppigerus.
 Fossil of Puppigerus parkinsonii, at Teylers Museum, Haarlem. Photo by Ghedoghedo Creative Commons Attribution, Wikipedia |
Range: Eocene of United States, England, Belgium, and Uzbekistan. (P. camperi is known from the London Clay and Bracklesham Beds of England, as well as the Sables de Bruxelles and the Sables de Wemmel of Belgium. P. nessovi is known from the Dzheroi 2 locality of Uzbekistan. P. crassicostata may or may not be the same as P. camperi).
Phylogeny: Cheloniidae : Euclastes + Gigantatypus + Argillochelys + Eochelone + Itilochelys + (Pacifichelys + (Cheloniinae + * ))
Comments: Another representative of the subfamily Eocheloniinae. From Wikipedia: Fossils show that Puppigerus was around 90 centimeters long, and its weight has been estimated as being somewhere around 9-14 kilograms. Although cheloniids such as Puppigerus first appeared during the Cretaceous, several traits of this genus give it more of a resemblance to modern cheloniids: its "huge" eyes pointed sideways rather than upward, unlike more primitive cheloniids, and its shell was completely ossified. The pygal (rearmost plate of the upper shell) also lacked a notch seen in earlier cheloniids. It was a herbivore, living off marine vegetation, and one of the "best-adapted" prehistoric turtles: its "unusually large" eyes helped it gather as much light as possible, and its specialized jaw structure kept it from accidentally breathing in water. Its front legs were flipper-like, but its hind legs were not developed in this manner, suggesting that it would have spent considerable time on dry land, where females would have laid their eggs.
 Fossil of "Chelonia" gwinneri, Wegner 1918, a small marine turtle from the Oligocene (Rupelien) of Flörsheim, Germany. Despite its good preservation, the phylogenetic placement of this species is unclear as it has not been determined cladistically. Many fossils of marine Cheloniids were traditionally identified as "Chelonia". Specimen at Senckenberg Museum of Frankfurt. Photo by Ghedoghedo Creative Commons Attribution, Wikipedia |
Cheloniinae : sea turtles (Green, Hawksbill, Loggerhead, Ridley, and their fossil relatives)
Range:
Phylogeny: Cheloniidae : Euclastes + Gigantatypus + Argillochelys + Eochelone + Itilochelys + (Pacifichelys + (Puppigerus + * : Syllomus + Chelonia + Natator + Carettini))
Comments: Included here are six species of extant sea turtles, Caretta caretta or Loggerhead, Chelonia mydas or green turtle, Eretmochelys imbricata or hawksbill, Natator depressus or flatback Turtle, Lepidochelys kempii or Kemp's Ridley turtle and the related Lepidochelys olivacea or Olive Ridley turtle, plus their fossil relatives. Most of these species are capable of hybridising with others in the family (Karl et al 1995) (see also essay on hybridisation among giant tortoises) The female returns to the same nest area every 1-3 years. Determing evolutionary relationships within the Cheloniidae on morphological grounds has been difficult. Much of the problems are due to overall similarity, with species and genera nearly indistinguishable in terms of external morphology (Bowen, 2003 p.8). The various species are distinguished by details such as the prefrontal scales on the head, the number of and shape of scutes on the carapace, and the type of inframarginal scutes on the plastron. Molecular phylogeny however provides a celare picture (Bowen, 2003), and we have followed here the recent consensus (Naro-Maciel et al 2008, Thomson and Shaffer 2010, Guillon et al 2012)
There are a number of early Cenozoic fossil forms that may or may not belong here. A large sea turtle from the Earliest Paleocene (Danian age) of Illinois has been assigned to the recent genus Chelonia (green sea turtle), but many such cases fossils assigned to living genera have proven to be mistaken or impossible to confirm, so this identification still requires corroboration (Jehle, 2006), and the very early stratigraphic date makes it dubious. The late Oligocene Carolinochelys, from the Chandler Bridge Formation of South Carolina has been suggested as a possible ancestor of the living loggerhead turtle Caretta caretta, although this has yet to be confirmed with cladistic analysis. A mounted skeleton at the Science Museum of Minnesota, in a swimming pose, is the first articulated specimen of this kind. It is about one meter long, and was found in association with whales, the dugong Crenatosiren, sea birds, the crocodile Gavialosuchus, other turtles, and fishes. (Anon 2009). The oldest undisputed cheloniids are an unnamed form from the Early Miocene (Brinkman 2009) of Oregan, and Procoplochelys grandaeva from the middle Miocene of eastern North America (Joyce et al 2004 p.1001).
 |
| Green turtle, Chelonia mydas, Gimp-Savvy.com, US National Oceanic and Atmospheric Administration, Public domain, via Wikimedia |
Range: Middle Miocene or Pliocene to Recent, Cosmop.
Phylogeny: Cheloniinae : Carettini + * : Chelonia + Natator + Syllomus
Comments: The clade Chelonia + Natator is supported by molecular phylogeny. Syllomus may be an early form. MAK130308
Syllomus aegyptiacus Lydekker, 1889
Range: .
Phylogeny: Chelonini : Chelonia + Natator + *
Comments: Syllomus is a pseudodont Sea Turtle widely distributed in the middle Miocene (Brinkman 2009 p.39), known from Egypt, Japan, the Atlantic seaboard (the Middle Miocene Calvert Formation of New Jersey and North-eastern Virginia, along with Procolpochelys and Psephophorus (Weems 1974, Anon 2009)), and possibly the Temblor Formation of California (Lynch and Parham 2003). It is clear that, like modern sea turtles, it had a ocean-wide distribution. This was a small sea turtle, with a plastron length of about 60 cm (Mlynarski 1976 p.48) Taxonomically it has been placed, with several similar forms such as Kurobechelys, in its own subfamily Syllominae (Bishop 2007). Although fossil material from Japan and North America is relatively complete material and cladistic studies done, the phylogeny placement remains ambiguous, the species being variously interpreted as a sister-taxon to extant Cheloniidae, a basal member of the crown group (Lynch and Parham 2003) or sister taxon of the Flatback Turtle Natator depressa (Parham & Pyenson 2010) and hence a member of the tribe Natorini. The genus is more primitive than extant cheloniids in that the canalis caroticus lateralis and base of the canalis caroticus internus were fully encased in bone (Brinkman 2009 p.45), although it is also possible that advanced features developed independently in several lines of cheloniinae. MAK130308
 |
| Green turtle, Chelonia mydas, Hawaii, Photo by Brocken Inaglory, GNU Free Documentation/Creative Commons Attribution license, via Wikimedia |
Range:Late Miocene to Recent, tropical and subtropical Oceans
Phylogeny: Chelonini : Natator + Syllomus + *
Characters: "Skull comparatively long, narrow, with the orbits lateral. Shell cordiform or pointed al both ends, with four costal shields, the plastral vacuities persisting for a long period. An intergular and a series of inframarginal shields present." Zittel, Eastman, et al (1902) pp.198. One pair of scales between eyes (Bishop 2007)
Comments: The extant green sea turtle, Chelonia mydas, has a range that extends throughout tropical and subtropical seas around the world. Although there are two distinct populations in the Atlantic and Pacific Oceans, these do not qualify as distinct subspecies. Adult green turtles grow to 1.5 metres in length with a weight of 68-190 kg and average carapace length is 78-112 cm, with exceptional specimens larger. Unlike other extant cheloniids,Chelonia is mostly a herbivorous, adults usually inhabit shallow lagoons, feeding mostly on various species of seagrasses. Like other sea turtles, green sea turtles migrate long distances between feeding grounds and hatching beaches. Many islands worldwide are known as Turtle Island due to green sea turtles nesting on their beaches. It is listed as endangered by the IUCN and CITES Wikipedia
 |
| Natator depressus, illustration from McCulloch 1908, public domain |
Natator depressa Flatback Turtle
Range: Recent, northern Australasia
Phylogeny: Chelonini : Syllomus + Chelonia + *
Comments: Unlike other sea turtles which have an ocean-wide distribution, the flatback sea turtle (Natator depressus) is endemic only to the continental shelf of northern Australasia. It usually found in bays, shallow, grassy waters, coral reefs, estuaries, and lagoons. The carapace of the adult is on average 90 cm long. It is low-domed, the edge is upturned, and has four pairs of costal scales—fewer than other marine turtles. The upper parts are an olive-grey, and more pale ventrally. A single pair of scales is located at the front of the head, which also distinguish this species. (Wikipedia). No fossils are known. Although initial molecular studies resolved this species as the sister taxon to the Carettini (Bowen, 2003, Iverson et al 2007), the current consensus is that it is more closely related to Chelonia mydas (Naro-Maciel et al 2008 Thomson and Shaffer 2010, Guillon et al 2012). It may or may not be related to, and presumably evolved from, the widespread Miocene Syllomus (Parham & Pyenson 2010)
Carettini Zangerl and Turnbull, 1955
Range: Early Miocene to Recent, Cosmop.
Phylogeny: Cheloniinae : Chelonini + * : Eretmochelys + (Procoplochelys + Caretta + Lepidochelys)
Characters: presence of two pairs of prefrontal scutes between eyes, four or more pairs of lateral or imbricated scutes
Comments: The clade Caretta + Lepidochelys is one of the few represented by morphological and molecular congruency. Procoplochelys would be an ancestral form from which these two evolved. The phylogenetic position of Eretmochelys is more uncertain, it may be a basal member of the Carettini, as indicated by molecular studies (Naro-Maciel et al 2008), or it could be related to Chelonia, as morphology suggests (Gaffney and Meylan 1988) MAK130307
 |
| Eretmochelys imbricata. The carapace's serrated margin and overlapping scutes are evident in this individual. Photo by Ecocentrik Guy, GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
Eretmochelys Hawksbill turtle
Range: Recent, Atlantic, Pacific, and Indian Oceans
Phylogeny: Carettini : (Procoplochelys + Caretta + Lepidochelys) + *
Characters: Four pairs of lateral or imbricated scutes
 |
| Hawksbill Turtle off the coast of New Ireland. A close-up of the hawksbill's distinct beak. Photo by Tom Doeppner, July 2000. More photos from that dive. GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
Comments: E. imbricata is easily distinguished from other sea turtles by its sharp, curving beak with prominent tomium. There is a distinct pattern of thick scutes that make up its carapace. While its carapace has five central scutes and four pairs of lateral scutes like several members of its family, E. imbricata's posterior scutes overlap in such a way as to give the rear margin of its carapace a serrated saw-like appearance of its shell margins. The hawksbill's forelimbs have two visible claws on each flipper. Hawksbill shells slightly change colors, depending on water temperature. While this turtle lives part of its life in the open ocean, it spends more time in shallow lagoons and coral reefs. Adult hawksbill sea turtles have been known to grow up to 1 meter in length, weighing around 80 kg on average. The turtle's shell, or carapace, has an amber background patterned with an irregular combination of light and dark streaks, with predominantly black and mottled-brown colors radiating to the sides. While this turtle lives part of its life in the open ocean, it spends more time in shallow lagoons and coral reefs. A critically endangered species, in the past its shell was used for tortoiseshell jewellry. (Wikipedia). We are here following molecular phylogeny in placing this species in the tribe Carettini.
Procoplochelys grandaeva (Leidy, 1851)
Range: middle Miocene of the eastern United States.
Phylogeny: Carettini : Eretmochelys + (Caretta + Lepidochelys + * )
Comments: Originally Chelonta grandaeva Leidy, the genus Procoplochelys was coined by Hay (1908) p.215 from then still fragmentary material. Procoplochelys grandaeva, from the middle Miocene Calvert Formation of New Jersey, seems to be closely related to the extant Lepidochelys and Caretta on the basis of features of the shell, particularly the increased number of neurals (see Zangerl and Turnbull, 1955) as well as cranial features Weems (1974). It can therefore be confidently placed in the Tribe Carettini, "Procoplochelys" melii (Misuri, 1910) from the Miocene Pietra Leccese sediments of south-eastern Italy is probably not related (Brinkman 2009 p.39)
| Caretta caretta (Loggerhead Sea Turtle) at Océanopolis, Brest, France. Photo by User:Strobilomyces , GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
Caretta Merrem, 1820 or Stejneger, 1904 : Loggerhead
Synonym: Thalassochelys caretta
Range: Late miocene/Pliocene to Recent, Cosmopolitan
Phylogeny: Carettini : Eretmochelys + (Procoplochelys + Lepidochelys + *)
Characters: "Carapace completely ossified in the adult, with at least five pairs of costal shields, and vacuities more or less obliterated. A series of inframarginal plastral shields present." Zittel, Eastman, et al (1902) pp.198
Comments: It's a shame that the old name (e.g. Zittel, Eastman, et al (1902) pp.198-9) Thalassochelys - "sea turtle" is no longer used. The loggerhead (Caretta), is a sea turtle with a cosmopolitan distribution. It first appears during the late Miocene or Pliocene (Parham & Pyenson 2010). The only well known fossil species is Caretta patriciae is a Pliocene species from North Carolina (Zug 2001). "Caretta" eocaenica Lydekker 1889 from the Eocene of England does not belong in this genus, but probably represents an unspecified stem cheloniid. C. caretta (Linnaeus, 1758) is the only extant species. There is no evidence that it should be divided into the two subspecies C. c. caretta and C. c. gigas (Bowen, 2003).
The following notes on the evolution and biogeography of the genus are from Wikipedia
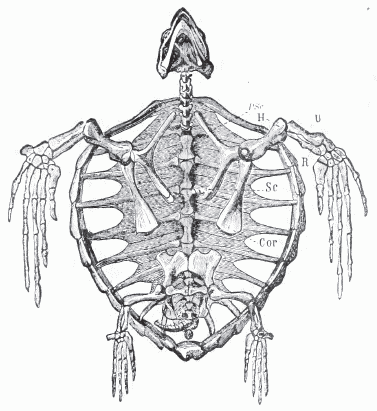 |
| Skeleton of the modern cheloniid Caretta caretta, Recent; Mediterranean, showing features of an advanced sea turtle, The lightweight and reduced shell, and the greatly enlarged fore-flippers for use in propulsion, with the smaller rear flippers used for steering, are feature that independently developed in all lineages of advanced sea turtles (including the unrelated Protostegidae) Ventral aspect of skeleton, the plastron removed, cor, Coracoid ; h, Humerus; pre, Precoracoid; r, Radius; sc, Scapula; u, Ulna.- Zittel, Eastman, et al (1932) p.199, public domain |
Around three million years ago, during the Pliocene epoch, Central America emerged from the sea, effectively cutting off currents between the Atlantic and Indo-Pacific Oceans. The rerouting of ocean currents led to climatic changes as the Earth entered a glacial cycle. Cold water upwelling around the Cape of Good Hope and reduction in water temperature at Cape Horn formed coldwater barriers to migrating turtles. The result was a complete isolation of the Atlantic and Pacific populations of loggerheads. During the most recent ice age, the beaches of southeastern North America were too cold for sea turtle eggs. As the Earth began to warm, loggerheads moved farther north, colonizing the northern beaches. Because of this, turtles nesting between North Carolina and northern Florida represent a different genetic population from those in southern Florida (Spotila, 2004 p.167).
The distinct populations of loggerheads have unique characteristics and genetic differences. For example, Mediterranean loggerheads are smaller, on average, than Atlantic Ocean loggerheads (Spotila, 2004 p.166). North Atlantic and Mediterranean loggerhead sea turtles are descendants of colonizing loggerheads from Tongaland, South Africa. South African loggerhead genes are still present in these populations today (Spotila, 2004 p.167).
Adults have an average weight range of 80 to 200 kg and a length range of 70 to 95 cm, although some specimens grow much larger (the maximum reported weight is 545 kg and the maximum carapace length is 213 cm). No external differences in gender are seen until the turtle becomes an adult, the most obvious difference being the adult males have thicker tails and shorter plastrons than the females. Wikipedia
Loggerheads can produce intergeneric crosses with most other members of the family Cheloniidae, even ones only distantly related. Molecular sequencing reeveals a long separation between Atlantic and Indian-Pacific populations, and regional clusters of genotypes in the western Pacific and South Atlantic, although this temperate adapted species can occasionally transcend geographic boundaries by transfer of mtDNA lineages between the Atlantic and Indian oceans via southern Africa. (Bowen, 2003)
 |
| Lepidochelys olivacea at Réunion. Photo by Thierry Caro, GNU Free Documentation/Creative Commons Attribution license, via Wikipedia |
 |
| Researchers collect Kemp's ridley (Lepidochelys kempii) sea turtle eggs. Washington DC Library. Obtained from http://images.fws.gov/. via Wikipedia, Public Domain |
Lepidochelys Fitzinger, 1843 : Ridley sea turtels
Range: Late miocene/Pliocene to Recent, Atlantic, Pacific, and Indian Oceans
Phylogeny: Carettini : Eretmochelys + (Procoplochelys + Caretta + *)
Characters: Five or more pairs of lateral scutes
Comments: Ridley sea turtles (Lepidochelys) are a genus of medium-sized sea turtles about 50-70 cm long. They feed on crabs, fish, cephalopods, clams, and some marine vegetation (Wikipedia). Both morphology-based cladistics and molecular sequencing agree that ridleys are very closely related to loggerheads, and both may have speciated from Procoplochelys during the late Miocene or early Pliocene (Parham & Pyenson 2010). There are two extant species, Lepidochelys olivacea or Olive Ridley turtle, and the slightly larger, rare and criticaly endangered Lepidochelys kempii or Kemp's Ridley turtle.
 |
| Leatherback turtle Dermochelys coriacea. Photo from SOS Turtles Panama. |
Dermochelyidae : leatherback turtles
Range: Late Cretaceous (Campanian) to Recent, Cosmopolitan
Phylogeny: Crown Group Chelonioidea : Pancheloniidae + * : Corsochelys + (Eosphargis + (Psephophorus + Dermochelys))
Comments: The family Dermochelyidae is represented today only by the leatherback turtle Dermochelys. This is a huge animal with a shell length up to 2 meters in length, 1 to 1.5 m in width, weighing up to 590 kg. As with the Cretaceous protostegids (with which dermochelyds have in the past been grouped), the leatherback turtle takes the reduction of the bony shell to extremes. The costalia dermal plates fused to flattened ribs and the neuralia (dermal plates over vertebrae) are greatly reduced, somewhat like in the soft-shelled trionychid turtles, and no longer fused to build a carapace. Instead of a hard shell and horny plates it bears a mosaic of numerous bony platelets situated in a thick, leathery skin external to the free ribs and vertebrae. The plastron is also greatly reduced, and at least in Tertiary specimens the ventral side shows secondary platelet armour.
Although the fossil record of the Dermochelyidae is poor, the family has a long evolutionary history, with the earliest undisputed dermochelyids occurring in the Late Cretaceous (Campanian) of North America (Parham and Stidham, 1999) and Japan (Hirayama and Hikida, 1998). The Dermochelyidae reached a peak of diversity during the Eocene period with at least six species worldwide (ref). Prominent fossil taxa include Psephophorus from the Eocene of Europe and Eosphargis from the Paleocene and Eocene genus of Europe and Eastern USA Joyce et al 2004, Jehle, 2006, Wikipedia
The unique morphology of Dermochelys coriacea has resulted in many early authors placing it in a monotypic genus (e.g., Dermochelys Blainville, 1816; Sphargis Merrem, 1820), family (e.g., Sphargidae Gray, 1825), or even suborder (e.g., Athecae Cope, 1871; Athecata Lydekker, 1889). Joyce et al 2004
The taxon Pandermochelys is used by Joyce et al 2004 to refer the clade uniting the Dermochelyidae and the Cretaceous marine Protostegidae, with "Dermochelyidae" applied to all marine turtles more closely related to Dermochelys coriacea than to Protostega gigas (the type species of Protostegidae). Subsequent cladistic analysis including the ancestral protostegid Santanachelys gaffneyi shows that the Protostegids are not Cheloniodea at all, but belong to more ancient and primitive lineage of stem-cryptodires or even stem testudines dating back to the late Jurassic (refs), and that the similarities to Dermochelyids are the result of convergence. Joyce et al 2004 p.1001 suggest that a new name Pandermochelys should be used as the stem taxon for Dermochelyidae, but as these two terms seem to be synbonymous we see little reason to prefer the former over the well established Dermochelyidae.
Range: Early Campanian Mooreville Chalk Formation of Alabama. A similiar form has been found in the Maastrichtian Ouled Abdoun phosphate basin, Morocco.
Phylogeny: Dermochelyidae : Dermochelyinae + *
Comments: Basal dermochelyid and the earliest known crown chelionoidean. As with other early sea-going forms like Santanachelys and Toxochelys, likely possessed developed, salt-excreting lachrymal glands. A primitrive feature in all three is a carapace with only minimally reduced coastal plates. Like the protostegid Archelon and the extant Dermachelys, Corsochelys had channels penetrating the subphyseal plate from bone into the cartilage, which means that Corsochelys would have reached their large size quickly through fast skeletal growth, as with the leatherback. (adapted from Wikipedia)
 |
| Adult Dermochelys coriacea, Leatherback Sea Turtle, public domain, via wikipedia |
Dermochelyinae : leatherback turtles: Cosmochelys, Dermochelys, Eosphargis, Psephoporus
Range: Paleocene or Early Eocene to Recent, Cosmopolitan
Phylogeny: Dermochelyidae : Corsochelys + * : + (Eosphargis + (Psephophorus + Dermochelys))
Description: "Carapace broken up into numerous mosaic-like pieces of dermal ossification, wholly unconnected with the vertebrae and ribs. Epidermal shields absent. Skull without descending parietal processes; ternpored region completely roofed, the scpiamosal joining the parietal. Humerus flattened ; limbs paddle-shaped, dateless, the digits of the manus much elongated ; phalanges without condyles" - Zittel, Eastman, et al (1902) pp.197
Comments: Following the taxonomy of Bishop 2007, Dermochelyinae can be used to refer to higher Dermochelyidae; equivalent to the Dermochelyidae of Zittel, Eastman, et al.

Range: Early Eocene London Clay and Ypresian of Belgium, possibly Paleocene (Thanetian Maryland/Virginia), and Eocene of Eastern USA
Phylogeny: Dermochelyidae : Corsochelys + ((Psephophorus + Dermochelys) + * )
Description: Carapace represented by a single median row of broad carinated scutes and a series of marginals on either side. Plastron probably devoid of tesserae. Skull broad and flat..- Zittel, Eastman, et al (1902) pp.197
Comments: Broad and flat shell 1 to 1.5m long, carapace reduced relative to cheloniids, palate as in Dermochelys. The Western European species Eosphargis gigas (Owen, 1861) has a skull upto 33 cm wide. (Zittel, Eastman, Woodward, et al(1932), p.315, Jehle, 2006)
Image: Eosphargis breineri skull cast at the Geological Museum in Copenhagen, photo by FunkMonk, GNU Free Documentation/Creative Commons Attribution, via wikipedia
Range: Eocene to Pliocene of Europe, Late Eocene of Egypt and Alabama, Miocene of Maryland (Zittel, Eastman, Woodward, et al(1932), p.316), Mid Eocene of New Zealand Kohler (1995
Phylogeny: Dermochelyidae : Corsochelys + (Eosphargis + (Dermochelys + *))
Description: "Skull shorter, thicker, and relatively larger than in Dermochelys; shell completely tessellated, sculptured ; scutes of the larger longitudinal row of the carapace devoid of carinae" - Zittel, Eastman, et al (1902) pp.197, (Zittel, Eastman, Woodward, et al(1932), p.316),.
Comments: Predecessor and most likely ancestor of the extant leatherback turtle.
 |
| Dermochelys coriacea, Leatherback Sea Turtle, from: amigosdomarnaescola.com, via Reptilis net |
Dermochelys coriacea (Vandellius, 1761). Leatherback turtle
Synonym: Sphargis
Range: Recent, Atlantic, Pacific, and Indian Oceans
Phylogeny: Dermochelyidae : Corsochelys + (Eosphargis + (Psephophorus + *))
Description: "Carapace completely, plastron incompletely bony in the adult, the former with seven, the latter with five keels ; plastral elements eight. Recent ; Atlantic, Pacific, and Indian Oceans. In this, the largest and sole surviving member of the family, the separation of the vertebral column from the carapace was deemed by Cope, Dollo, and others a character of sufficient importance to warrant the establishment of a group (Athecae) equal in value to the rest of the Chelonia. Baur has endeavoured to prove that the mosaic-like carapace was an acquired character, and that this and the following family are directly related to the Chelonidae. - Zittel, Eastman, et al (1902) pp.197
Comments: largest living turtle (more), and the only surviving representative of an ancient family. As a highly advanced Dermochelyid, it has a reduced, soft, cartilaginous shell without scutes, , a throat and upper digestive tract lined with spines (to stop swallowed jellyfish from escaping) and an ability to generate some of its body heat from within. (more)
Page last modified MAK110922. All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners
{kind=link}
{kind=link}
{kind=link}
{kind=link}