|
|
Chelonii |
| The Vertebrates |
The mystery of Chelonian origins |
Chelonii - Odontochelys
Taxa on This Page
- Chelonii
- Odontochelys X
The Mystery of Chelonian Origins
|
This
|
or
|
this?
|
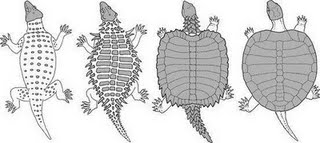
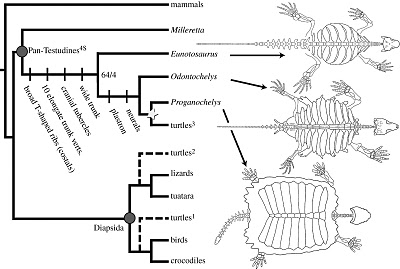
Above: Hypothetical sequence of events leading to the modern chelonian shell, in which dermal scutes expand to form the carapace (upper shell). The two animals on the left are speculative. The third one from the left is Proganochelys, and the far right is Chinlechelys. This sequence supports the theory that chelonians evolved from a procolophonid or pareiasaur-like ancestor (Joyce & Gauthier, 2004)
Right: the proto-chelonian Odontochelys. This animal would seem to support the idea that turtles evolved from aquatic, sauropterygian-like ancestors.
|
|

|
Graphics from "A Carapaceless Turtle" link/link(mirror postings)
The origin of turtles and tortoises (Chelonia) from ancestral reptiles is still unclear. There are many different rival theories of where Chelonians fit on the evolutionary tree, each supported by various arguments and with each there are also objections. "Turtles are very derived. Inevitably, they have convergently evolved derived characters with other groups. Which hypothesis we trust depends on which characters we value." - John Merck. . These hypotheses can be summarised briefly as follows:
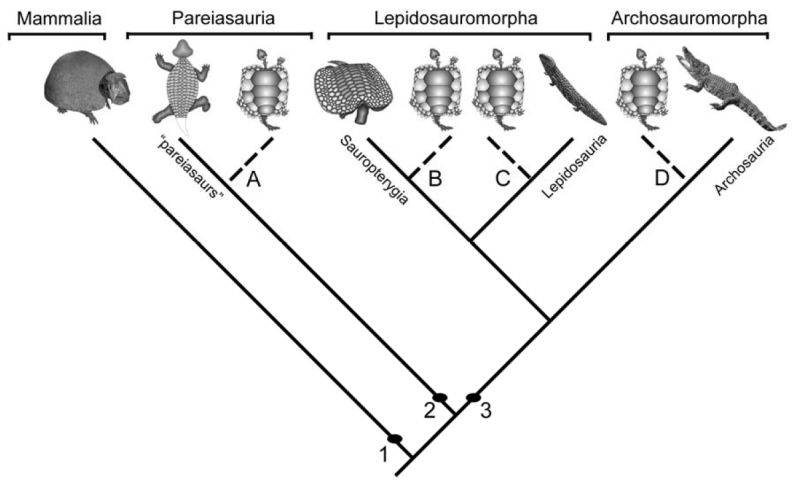
A few alternative chelonian origins: hypothesis A) chelonians are parareptiles and deeply nested within pareiasaurs; hypothesis B) chelonians are derived diapsids most closely related to sauropterygians; hypothesis C) chelonians are derived diapsids most closely related to lepidosaurs; hypothesis D) chelonians are derived diapsids most closely related to archosaurs. For source of data on phylogenetic hypotheses of chelonians see text. 1= Synapsida; 2= Parareptilia; 3= Diapsida. Diagram from Scheyer 2007 p.10
Eunotosaurs: The strange Permian reptile Eunotosaurus africanus was described by D. M. S. Watson as the ancestor to chelonia, on the basis of the broadly expanded ribs which form a shell-like structure covering its body (Watson, 1914). It was since discovered that the shell of chelonians is made up of narrow ribs covered by dermal bone, not broadly expanded ribs. This, plus the presence of an ectopterygoid bone in the eunotosaur skull, indicate that this creature was not a chelonian ancestor: Interestingly the Eunotosaur ancestry argument has recently been revived (Lyson et al 2010, see illustration at right). The authors suggest that because the ribs of Eunotosaurus were not only expanded, but T-shaped in cross section, this suggests that expansion occurred through secondary ossification and the incorporation of dermal (skin armour) components as in true chelonians, which answers the broad rib objection. Unfortunately, this still does not address the "evo-devo" model of the embryonic formation of the chelonian shell and the location of the shoulder girdle inside the ribcage, which is supported by Odontochelys (Willmore, 2010). We can safely say therefore that eunotosaurs developed a shell quite independently of chelonians. (also forum discussion of this paper)
Captorhinids: A more common view until fairly recently was that the closest extinct relatives of chelonians were the primitive Permian "cotylosaurian" reptiles of the family, Captorhinidae. Like all chelonians, members of this group lack the ectopterygoid bone and have a large medial process of the jugal bone of the skull. Gauthier et al. (1988) argue strongly for close affinities between captorhinids and chelonians. However, no known captorhinid shows any sign of even a partly developed shell. There is also a gap of many millions of years between the last known captorhinid and the earliest known chelonian. This idea has now been pretty much abandoned.
Procolophonids: During the early 90s, it was proposed by Robert Reisz and one of his students Michel Laurin that the Triassic herbivorous anapsids called Procolophonids are the closest relatives of chelonians. A study by (Reisz & Laurin 1991 of the basal procolophonoid Owenetta, revealed a number of characters unique to procolophonoids and chelonians (see Procolophonoidea: Morphology). Most notable is the tympanic notch (the ear). Chelonians lack a tympanum (a thin membrane like the skin of a drum that enables the reception of airborne vibrations, hence, "ear drum") but instead have a layer of skin over their tympanic notch or opening. This unique arrangement is alos found in Owenetta, but not in other reptiles. However, there is again a gap in time (although not as great as with the Captorhinids between Owenetta and its relatives and the chelonia. Once again, no known procolophonid shows any tendency of even a partly developed shell.
Pareiasaurs:  . Beginning in 1993 (Lee 1993), Michael Lee has argued that the large armoured herbivorous Permian pareiasaurs are the closest known relatives of chelonians, rather than the small procolophonids proposed Pareiasaurs as a sister taxon. Pareiasaurs and chelonians share a number of traits, including a thickened braincase floor, acromian process, dermal armour, a shortened vertebral column and four presacral ribs, among many other features. Cladistically speaking, chelonians are therefore a type of specialised dwarf pareirasaur that survived the end Permian extinction (Lee 1996; Lee 1997). This is the same as birds as specialised dinosaurs. Lee coined the wonderful term Hallucinocrania to refer to the clade that includes pareiasaurs and chelonians (with reference to the bizarre form of the pareiasaur skull bones) This theory, which became for a while the established one, is supported by the fact that towards the end of the Permian some pareiasaurs became reduced in size, and some, such as the smallish genus Anthodon (left) developed a chelonian-like pattern of bony plates from the dermal armour (osteoderms) embedded in their skin, forming a rigid covering over the entire back, and of flattened, expanded ribs. The discovery of Chinlechelys (Joyce, et. al. 2008) supports the argument for an osteoderm-bearing (pareirasaur-like) terrestrial ancestor.
. Beginning in 1993 (Lee 1993), Michael Lee has argued that the large armoured herbivorous Permian pareiasaurs are the closest known relatives of chelonians, rather than the small procolophonids proposed Pareiasaurs as a sister taxon. Pareiasaurs and chelonians share a number of traits, including a thickened braincase floor, acromian process, dermal armour, a shortened vertebral column and four presacral ribs, among many other features. Cladistically speaking, chelonians are therefore a type of specialised dwarf pareirasaur that survived the end Permian extinction (Lee 1996; Lee 1997). This is the same as birds as specialised dinosaurs. Lee coined the wonderful term Hallucinocrania to refer to the clade that includes pareiasaurs and chelonians (with reference to the bizarre form of the pareiasaur skull bones) This theory, which became for a while the established one, is supported by the fact that towards the end of the Permian some pareiasaurs became reduced in size, and some, such as the smallish genus Anthodon (left) developed a chelonian-like pattern of bony plates from the dermal armour (osteoderms) embedded in their skin, forming a rigid covering over the entire back, and of flattened, expanded ribs. The discovery of Chinlechelys (Joyce, et. al. 2008) supports the argument for an osteoderm-bearing (pareirasaur-like) terrestrial ancestor.
Despite the plausablity of this hypothesis, it is not without problems. The biggest problem is that chelonian carapace ('dermal armour') does not develop from dermal plates in the skin as with pareieasaurs. It, is a complex structure formed through fusion of vertebrae and ribs to interlocking plates beneath the skin and growth of horny scutes above the skin (Cherepanov 1997; Rieppel and Reisz 1999), without parallel in the animal kingdom. Moroever, the ancestral chelonian could not have been a terrestrial animal. The unique chelonian morphology, in which the limb girdles and shoulder blades develop within the rib-cage, rather than outside as in all other vertebrates, mean that the ribs would somehow have had to lose their function of supporting respiration and locomotion Rieppel and Reisz 1999), and hence implies an aquatic ancestry, exactly what we find with Odontochelys. Hence the Sauropterygia (Triassic aquatic) origin theory.
There are more technical objections as well. Derived chelonians have lost the basipterygoid joint, as have pareiasaurs, but early forms such as Odontochelys and Proganochelys retain it. Odontochelys teeth are simple cones, very different from the leaf-shaped teeth of pareiasaurs (David Marjanovic DML post). Then there is the recent discovery of what seem to be cleithra in the stem chelonian Kayentachelys, and even stem-pleurodires and stem-cryptodires (Joyce et al 2006) (I am not sure if it has been identidfied in Odontochelys). Cleithra are a primitive element of the shoulder girdle that are present in stem synapsids and many anapsid reptiles, but absent in all diapsids, as well as in pareiasaurs. This favours a parareptile ancestry, but not a pareiasaur or diapsid origin.
Sauropterygia: The turtles and placodonts (aquatic animals with broad wide shells) show superficial similarities in anatomy, and according to Broom that chelonians and placodonts were related, although in many morphological respects they are very different. This idea has recently been revived by Olivier Rieppel (Rieppel 1995, Rieppel and deBragga 1996, deBraga and Rieppel 1997). Rieppel and deBraga pointed out that previous phylogenetic studies had been too narrowly focused, being based on the assumption that turtles were anapsids. Once they broadened their analysis to include other groups they discovered, that testudines nested within the diapsids, and more specifically inside the Euryapsida (placodonts and sauropterygia), a group of marine reptiles that was established by Romer. Shared features include jaw adductor muscles, temporal (cheek) bone ossification, the shape of the jugal, and the presence of a centrale among the carpal (wrist) bones instead of a radiale (forarm). This was the first cladistic analysis to move chelonians out of their until then universal position within the Anapsida, to the diapsids. This means that chelonians are not derived from an extinct anapsid group but diapsids (and more specifically lepidosauromorphs, as the authors consider sauropterygia and lepidosauria sister groups) that secondarily closed their temporal fenestrae and lost their impedance-matching ear in reverting to the basal amniote condition. Synapomorphies include a hooked fifth metatarsal and an astragalus-calcaneum complex (Lyson & Gilbert 2009). They found that testudines nested particularly close, and even within the sauropterygia, especially placodonts. Rieppel and Reisz 1999 hypothesized a marine origin for the ancestral chelonian, that it most likely had dorsally located ribs, and the plastron developed first in order to protect the soft underbelly. And while the plastron might be derived from gastralia, the sauropterygian pectoral and pelvic girdles form expanded, nearly flat plates meeting along the ventral midline, and this could be another way in which the plastron developed (blog comment). Because Sauropterygia are common and well established in the Triassic, it would certainly explain why chelonians are unknown in the fossil record until the late Triassic.
The discovery of Odontochelys adds weight to the Sauropterygia origin theory (Li et al 2008; Reisz & Head 2008), although this strange form also arises more questions than it answers , and it could equally be used to argue for a parareptilian ancestry, either eunotosaur (Lyson et al 2010) or procolophonoid (David Marjanovic blog comment ). In any case even in this very primitive turtle there is no trace of temporal fenestration such as would characterise it as a diapsid, in contrast to all other specialised aquatic Triassic reptiles.
There is a problem however with placodonts. Although extremely turtle-like, they developed their shell by fusing osteoderms - bony armor plates that develop from the skin. Whereas the chelonian shell is a unique combination of dermal and endochondral (skeleton) bone. So the placodont connection has the same problem the pareirasaur one has. In each case, the structure, and indeed the microstructure of the armour is quite different to turtles and tortoises (Scheyer 2007). And whilst Priscochelys hegnabrunnensis, based on a fragmentary piece of shell from the Muschelkalk of Germany was originally believed to represent the most ancient chelonian, and a transitional form between placodonts and testudines (Karl, 2005), it was later shown to be a cyamodontoid placodont (Scheyer 2008).
The analysis used has been criticised (Lee, 1997a), and the sauropterygian-chelonian theory is only weakly supported and based on characters that are homoplastic on the tree as a whole (Rieppel and Reisz, 1999), with only a small increase in tree length needed to return turtles to the Parareptilia (Wilkinson et al. 1997). There is also the matter of taxon sampling. Turtles appear as diapsids, and the sister clade to, or even nested within, the sauropterygians in the analyses including euryapsids; without euryapsids, the same analysis supports a parareptile affinity for turtles (Brochu 2001, pp.1190, 1192), while adding additional characters and taxa mak them the sister group of the lepidosaurs (Hill, 2005).
One thing Odontochelys does do is conclusively refute the pariesaurian hypothesis. Quoting Adam Yates (Closing in on turtle origins):
Of the synapomorphies linking pareiasaurs, or derived subclades within pareiasaurs, to turtles a good many of them are missing in Odonotochelys. These include: basal tuberae (ventral swelling of the braincase) midway between the occipital condyle and the basipterygoid processes (where the palate attaches to the braincase); acromial process on the scapula, closure of the spaces between the ribs (it is debatable wether or not any turtle actually has this condition); fewer than twenty caudal vertebrae; and body covered in united osteoderms. Note that although it appears to support a diapsid origin for turtles the skull of Odontochelys lacks any trace of temporal openings so perhaps we can't quite rule out other anapsid sister groups just yet.
Lepidosaurs: although there are no transitional forms known between the lizard and lizard-like lepidosaurs (a group that includes lizards and snakes and the sphenodontia (tuatura), and seems to be allied to the Sauropteerygia) and testudines, there is a decent amount of cladistic evidence that argues for their close relationship. Of course, these may just be parallels, but the lepidosaur origin, while not as popular as the pareiasaur, sauropterygian, or now archosaur orgin, still has supporters and cladistic evidence (Müller, 2003.; Hill, 2005, the latter, using much larger taxonomic and character sampling, arriving at the rather surprising finding that Lepidosaurs and Crown group Archosaurs (to the exclusion of basal archosauromorphs) are also sister taxa, which goes against the entire Diapsid phylogenetic consensus).
Archosaurs. Yes, archosaurs! Beginning in 1997, molecular phylogenetic analyses, both mitochondrial and nuclear gene analyses, consistently and unanimously places suggests that testudines are related to archosaurs, or even that they are derived archosaurs. (Zardoya and Meyer 1998; Hedges & Poling, 1999 (review); Kumazawa & Nishida, 1999; Janke et al. 2001; Zardoya and Meyer 2001; Meyer and Zardoya 2003; Rest et al 2003; Hugall et al. 2007; Shedlock & Edwards, 2009). Although one early findings (of Hedges & Poling, 1999) rather disconcertingly also placed Sphenodon (the tuatura) among the archosaurs, this was not reported in the later paper by Rest et al 2003, who however confirm the testudine-archosaur sister relationship.
This unanimous molecular evidence may or may not be the result of factors such as long branch attraction, always a problem with isolated taxa - here molecular evidence for archosaurian affinities of turtles is based on ingroup divergences little older than the Jurassic, whilst available outgroups (mammals and lissamphibians) had diverged by the end of the early Carboniferous (Brochu 2001, p.1190). - but it is unanimous across all forms of molecular analysis (Shedlock & Edwards, 2009 p.376). And while early molecular analyses could be criticized (short sequences, poor taxon sampling, genes with inappropriately fast substitution rates), and molecular phylogeny prior to 2001 tends to suffer from long-branch attraction (citing David Marjanovic post), more recent analyses of long nuclear genes such as the nuclear gene RAG-1 ((Recombination activating gene, a large slowly evolving gene useful for phylogenetic analysis (Krenz et al 2005, p.186.)) still strongly position chelonians as the sister group to archosaurs (Hugall et al. 2007). Although the much slower nucleotide substitution rate in turtles in relation to diapsids might predispose the reptile RAG-1 tree to be rooted in diapsids, there is no difference in substitution rate in the case of mitochondrial genes Rest et al 2003 and studies here also place testudines alongside archosaurs (Lyson & Gilbert 2009). The only disagreement being whether chelonians are closer to birds or to crocodiles, or equally distant from either. It has even been suggested that aëtosaurs are stem-turtles; advanced forms like Typothorax and Paratypothorax had broad turtle-like bodies, their ventral plating compared to a plastron, and in at least one species the neck spines resemble those of an early turtle (Hedges & Poling, 1999). But these are quite weak arguments, much better explained as homoplasy, and if chelonians do belong on the archosaurian tree it is more likely as basal Archosauromorphs. In any case when both crocodiles and birds are used as outgroups to test intra-testudine phylogeny they come up as more closely related to each other than to testudines (e.g. Krenz et al 2005, see figure here). The problem is, this hypotheses has virtually no morphologic support, and is equally opposed by both the Parareptile origin and Sauropterygian origin camps (Rieppel 2000; Lee, 2001); and it has been argued that even when both morphology and molecules are considered together a parareptile ancestry is still the most plausible (Lee, 2001), There are more recently a few morphological arguments. Comparison of chelonian bone histology with osteoderm-bearing pareiasaurs, mammals, placodonts, archosauromorphs and lepidosaurs, show closest resemblance with archosaurs (Scheyer 2007). And a new paper reports the existence of an archosaurian-like laterosphenoid bone in Proganochelys (Bhullar & Bever 2009). There is also analysis that supports Ichthyosauria, Sauropterygia and Chelonia included under the Archosauromorphs (but not Archosauria) rather than Lepidosaurs ( Merck, 1997, Borsuk-Białynicka & Evans 2009 p.230), which would be an interesting way to converge the morphological (Rieppel & Braga's Sauropterygia + Chelonia) and molecular findings. See also Azendohsaurus, the early archosauromorph radiation and a new position for turtles for an interesting if speculative approach to this, placing testudine ancestry within the Archosauriforms. And it is worth considering the quite similar problem of the origin of whales, another highly specialised group that did not have clear terrestrial ancestors. Against all expectations, molecular phylogeny nested Cetacea (whales and their relatives) with the Artiodactyla (even-toed hooved animals). This was confirmed by a discovery in 2000 of the distinctive artiodactyl-like double-pulley astragalus (ankle) bones in early archaeocete skeletons (Gingerich et al., 2001, non-technical essay) There are however other isntances where molecular evolution gives much more dubious results, such as grouping monotremes and marsupials together, the Marsupionta hypothesis (Janke et al. 1996). However, unlike the case with turtles, these findings were alter challenged by further molecular research (e.g. Killian et al. 2001; Kullberg et al 2008)
Yet while both cladistic morphological, and molecular phylogeny have been widely employed, and as above mentioned give very different results, a third methodology, embryology, was brought in by Werneburg & Sánchez-Villagra 2009 to examine and test these conflicting phylogenetic hypotheses. They found that the timing of organogenesis events (stages of embryonic development) in the various amniote clades clearly supported a basal position of turtles (i.e. as suister group to Sauria). This implies significant convergence or paralellism in either the molecular/genetic composition, adult morphology, or embryological developmental timing characters, as only one of these alternative solutions can be right. The following presents
Thus the problem remains unresolved, and the conficting arguments, each persuasive in its own way, make it impossible to identify any ancestral group with confidence. The Eunotosaur and Pareiasaur hypotheses can I believe be discarded. As for the rest, molecular phylogeny and shell histology make a powerful case for the Archosauromorph origin, and gross morphology equally supports anapsid/parareptile and lepidosauromorph origins. But embryology, the presence of cleithra in basal and stem chelonians, and the absense of any trace of temporal openings (on its own not very persuuasive because temporal fenestrae disappear in many diapsid lineages, but perhaps significant in association with the other arguments) even in such a transitional form as Odontochelys (the chelonian equivalent of archaeopteryx if ever there was one), all seem to suggest an anapsid ancestor, perhaps a hypothetical aquatic Owenettid-type procolophonoid. Admittedly, the lack of any evidence of such an animal in the fossil record does make this hypothesis a lot less plausible than it otherwise would be. (MAK101013)
Some Links/References used here: John Merck - class notes (On Beyond Dinosaurs: Patterns and Enigmas in Vertebrate Evolution) (includes very good description of Long branch attraction) (updated page with Odontochelys, but some nonloading images). Reptipage - T-U-R-T-L-E Power! Part 2: The weird and wacky origin of turtles (easy to read page, some of this information included here). Turtles all the way down: loggerheads at the root of the chelonian tree - pdf - short and readable academic essay) MAK101030
 The proto-chelonian Odontochelys semitestacea - Carnian of China
The proto-chelonian Odontochelys semitestacea - Carnian of China
Artworkby Arthur Weasley - Wikipedia
Descriptions
Chelonii Latreille 1800 : ancestral and derived turtles (= Testudinata).
Axial arrest of ribs, plastron. For derived forms see Testudinata.
Notes: Usually Chelonia, Testudines, or Testudinata. However Dubois & Bour 2010 argue that none of these names are viable. For example, although "Testudines" has been credited to Linnaeus, the term as originally used was a plural referring to the members of the genus Testudo, not a family-series or class-series nomen. Both Chelonii and Chelonia are latinisations of Chéloniens Brongniart, 1800. Chelonia was first used in 1802, after Chelonii (1800), and moreover can lead to confusion because of homonymy with the generic name Chelonia Brongniart, 1800. Testudinata is a valid name coined by Oppel, in 1811, Chelonii of Latreille, which dates to 1800, has priority over the former, as well as over Chelonia Ross & Macartney, 1802 (Dubois & Bour 2010, pp. 152-3). Testudines and Chelonia(ns) can still be used as informal names. MAK121222
Odontochelys: most primitive known chelonian. O. semitestacea Li et al., 2008
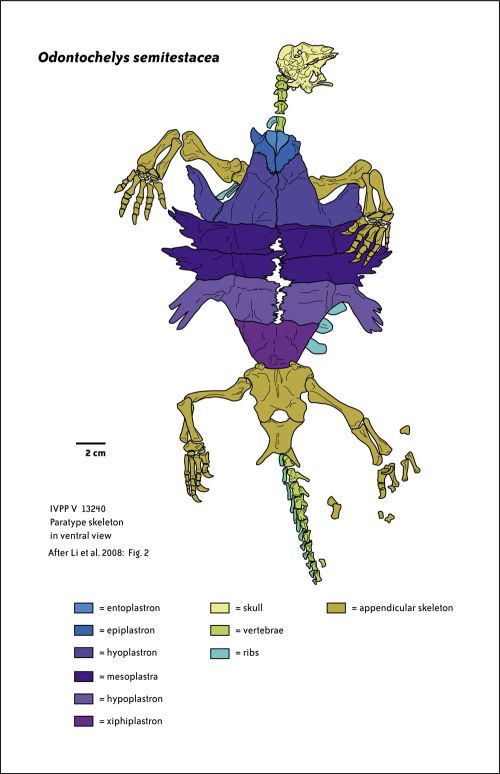
Description and Comments: from Wikipedia, slightly edited: Odontochelys semitestacea ("toothed turtle with a half-shell") is the oldest known chelonian. It is the only known species in the genus Odontochelys and the family Odontochelyidae. The species was first described from three 220 million year-old specimens excavated in Triassic deposits originating from Guizhou, China. (Li et al., 2008; Reisz & Head 2008)
As a primitive chelonian, Odontochelys grossly differed from modern representatives of the group in several ways. Modern chelonians possess a horny beak without any teeth in their mouth. In contrast, Odontochelys fossils were found to have had teeth embedded in their upper and lower jaws. One of the most striking features of chelonians, both modern and prehistoric alike, are their dorsal shells, forming an armored carapace over the body of the animal. Odontochelys only possessed the bottom portion of a turtle's armor, the plastron. It did not yet have a solid carapace as most other testudines do. Instead of a solid carapace, Odontochelys possessed broadened ribs like those of modern chelonian embryos that still have not started developing the ossified plates of a carapace. (Li et al., 2008)
Aside from the presence of teeth and the absence of a carapace, a few other skeletal traits distinguish Odontochelys as basal compared to other chelonians, extant and otherwise. The point of articulation between the dorsal ribs and the vertebrae are decidedly different in Odontochelys than in later testudines. In a comparison of skull proportions, the skull of Odontochelys is far more elongated pre-orbitally (behind the eyes) compared to other chelonians. The tail was proportionally long, and unlike later forms, transverse processes in the tail are not fused. The scapulae of the examined specimens were identified to lack acromion processes. These anatomical differences are all very primitive features, indicating the transitional nature of this species (Li et al., 2008).
It is likely that Odontochelys was aquatic, as the fossil specimens were found in marine deposits rife with conodonts and ammonites. It probbaly frequented shallow marine waters close to shore. (Li et al., 2008). In contrast, all other early chelonians were terrestrial until the Mid Jurassic. The aquaticx or semi-aquatic nature of Odontochelys supports a Sauropterygian origin for the group
The following is paraphrased and slightly modified from Mysteries of the Turtle's Shell (Nature Notes):
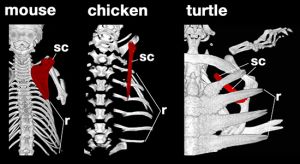 |
| Position of scapula (red) and ribs (r) in a mouse, chicken, and turtle. Note that in the turtle the scapula sits under the ribs, the only instance of this in the animal kingdom. In Odontochelys the scapula occupies an intermediate position, and is not uyet positioned inside the ribcage. Photo © Jamie Wilson / Shutterstock |
Turtles are unique not only in the way the shell is formed by the ribs being flattened and fused to the vertebrae, but even more extraordinarily by the placement of their shoulder blades (scapula). Rather than being located on the back, as in other vertebrates, they lie within its ribcage (see image, left). This results in the muscles connecting the shoulder blades to the trunk being twisted and folded.
For a long time scientists puzzled over how this bizarre arrangement could have evolved. Until recently, no one really knew. No transitional fossil forms had been found that showed the shoulder blade migrating towards the inside of the ribcage. In the end the new science of developmental biology, or "evo-devo" provided the answer. A team of scientists led by Hiroshi Nagashima from the RIKEN Center for Developmental Biology in Japan compared the embryos of turtles at various stages of development to those of mice and chickens ( Nagashima et al. 2009. During the early stages of development, all three embryos have shoulder blades that sit on their back, outside of the ribcage. As the mouse and chicken embryos develop, the shoulder blades remain on the animal's back, while the ribs become embedded in a layer of muscle called the muscle plate. But as the turtle embryo develops, the second pair of ribs swings forward and grows over the shoulder blades, while the muscle plate tucks inward and forms the edge of the turtle's shell.
Serendipitously, Ngashima's work was confirmed by the discovery of Odontochelys. Onontochelys lacks an upper shell (carapace) but posesses a lower shell (plastron). It has short ribs that did not fan out as with turtles proper. This means that Onontochelys' second pair of ribs did not swing over its shoulder blades during its early development. A veritable chelonian Archaeopteryx, Onontochelys is the long-awaited transitional form that also confirmns the evo-devo model, and vice-versa. MAK130121
Links: Figures and Tables from the above-referenced article ''An ancestral turtle from the Late Triassic of southwestern China'' in the Journal Nature; Oldest Turtle Found; May Crack Shell-Evolution Mystery; First known turtle swam on the half shell; Hairy Museum of Natural History; Niches - Turtleology MAK100520
Image: Hairy Museum of Natural History - larger image
Page last modified MAK110922. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners
{kind=link}