|
|
Chelonii |
| The Vertebrates |
Cryptodira: Paracryptodira |
Chelonii: Cryptodira: Paracryptodira
Taxa on This Page
- Arundelemys
- Baena
- Baenidae
- Baenini
- Baenodda
- Boremys
- Cedrobaena
- Chisternon
- Compsemys
- Dorsetochelys
- Eubaena-Stygiochelys lineage
- Glyptops
- Palatobaena
- Paracryptodira
- Peckemys
- Plesiobaena
- Plesiobaena-Palatobaena lineage
- Pleurosternidae
- Pleurosternon
- Stygiochelys
- Trinitichelys
Descriptions
Paracryptodira
Range: Jurassic to Eocene, Laurasia
Phylogeny: Testudines crown group : Pleurodira + Eucryptodira + * : Compsemys + ((Dorsetochelys + Pleurosternidae) + (Arundelemys + Baenidae))
Characters: reduced prefrontal exposure on the dorsal surface of the skull, reduced fenestra perilymphatica, secondary reduced supraoccipital crest. posterior foramen for the internal carotid canal located midway along the basisphenoid-pterygoid suture. (Gaffney, 1975 via Wikipedia)
Comments [1] Compared to more basal turtles, these forms have a more advanced system of blood flow to the head and a simplified plastron (Meylan 2001), although they are still more primitive than living cryptodires in the arrangement of the carotid arteries (Carroll 1988) and were unable to withdraw their head into their shell. MAK121118
[2] The discovery of the clade Paracryptodira is thanks to the early cladistic analyses of Eugene Gaffney (Gaffney, 1975 and later papers); prior to that time paracryptodires were included in the generic stem-group Amphichelydia, a testudine equivalent of cotylodsaurs, thecodonts, and condylarths. Gaffney's research, using Hennigian methodology that emphasised a small number of well understood synapomorphies, showed that the Paracryptodires were the sister group to the Eucryptodires (all recent cryptodiran turtles and their fossil relatives). The later change of methodology to statistical-computational trees unsettled this finding somewhat. A phenomenon first noted by (Lipka et al 2006 p.305, is the tendency of statistical trees to resolve Paracryptodira as stem pleurodires (see also for example fig and fig). This may be explained in terms of shared primitive characteristics, which wouyld be the reason why some paracryptodires such as Pleurosternon were previously classified as pleurodires (e.g. Zittel, Eastman, et al (1902)). Molecular phylogeny and stratigraphy both indicate that pleurodires, paracryptodires, and eucryptodires all emerged at around the same time from mid Jurassic stem testudines (Danilov & Parham, 2006). Since pleurodfires are a specialised offshoot of the primitive "cryptodire" condition (see Joyce 2007 and subsequent cladistic analyses; note that these authors limit the term "cryptodire" to the crown group only, contra Gahhney and co-workers) it is difficult to determine the exact branching point, and we have shown these groups as an unresolved trichotomy (the situation is further complicated by some cladistic analyses (e.g. Sterli 2008 and 2010, Pérez-García & Murelaga (2012)) not recognising Eucrptodira) MAK130311
Links: Wikipedia.
Compsemys Leidy.
Range: Late Cretaceous (Campanian) and Paleocene of N Am
Phylogeny: Paracryptodira : ((Dorsetochelys + Pleurosternidae) + (Arundelemys + Baenidae)) + *
Description: "Nasals free; a squamoso-parietal arch; descending processes of prefrontals joining vomer; stapes in an open groove of the quadrate; pterygoids narrow in the middle, without ring-like lateral expansions, separating quadrate and basisphenoid ; epipterygoid well developed and free; dentary bones distinct. Cervical vertebrae with well -developed transverse processes, with single articular faces, biconcave; dorsal and surra! vertebrae with well -developed ribs. Pelvis not anchylosed to the carapace and plastron. Epiplastra in contact with hyoplastra, entoplastron oval or rhomboidal; a complete series of marginals connected with the ribs." (Zittel, Eastman, et al (1902), p.206)
Comments: A common but poorly known genus, the basis for the paraphyletic suborder Amphichelydia Lydekker. Cladistic analysis places it as a basal paracryptodire (Lyson & Joyce 2011). The basal placement of Compsemys in the Paracryptodira implies a minimum ghost range of 70 Ma, from the Tithonian to the Campanian. But as the shell sculpturing of the most common Morrison turtle, Glyptops plicatulus, is almost identical, it is quite possible that many shell fragments attributed to G. plicatulus in particular, or Glyptops in general, may actually belong to Compsemys. Ironically, G. plicatulus was originally named Compsemys plicatulus by Cope, 1877. This would make Glyptops a junior synonym of Compsemys, and that either the species currently included in Glyptops consist of several distinct and even unrelated lineages (some of which would require new generic names), or that Compsemys victa is actually a highly derived pleurosternid. This latter, non-parsimonious (according to the cladograms by Lyson and co-workers) hypothesis is in keeping with the general tendency of molecular phylogeny to reveal morphological evolution to be endemically homoplastic and characteristred by frequent reversals (or, alternatively, for morphology to reveal the same about macromolecular evolution; the current consensus in phylogenetics however is to for the former). Compsemys' lack of specialised paracryptodire synapomorphies may also be compared with the similar situation in the Chelydridea, resulting in its basal placement in Hennigian cladistics, in contrast to the highly derived position given by molecular phylogeny.
There are a number of striking similarities between C. victa and the equally enigmatic extant big-headed turtle Platysternon megacephalum (either a chgelydrid or a testudinoid, depending on your choicxe of phylogenetic methodology), which would be the result of convergences due due to similar environments and carnivorous diet, hence the large head and well-developed premaxillary beak.
Based on the fine grained lithology in which it is most commonly preserved, Compsemys victa has been interpreted as inhabiting slow moving or ponded water environments. Taphonomic evidence reveals that other basal paracryptodires had similar environmental preferences. The quarries from which much of the Morrison (Kimmeridgian-Tithonian) possible pleurosternids Dinochelys whitei Gaffney, 1979, Uluops uluops Bakker, 1990, and G. plicatulus material was recovered (Dry Mesa Quarry, Breakfast Bench Quarry, and Quarry Nine, respectively) are thought to represent low energy pond or swamp environments, indicating that this is the original paleoecology for North American paracryptodires. (Lyson & Joyce 2011), in contrats to the lagoonal-coastal Pleurosternon, clearly a specialised ecomorph
Lyson & Joyce place all Campanian to Paleocene specimens in the single species Compsemys victa, which indicates either a very long-lived species or the possibility of this being divided into chrono-species. MAK130407
Dorsetochelys
Phylogeny: Paracryptodira : Compsemys + ((Arundelemys + Baenidae) + (Pleurosternidae + *))
Range: Late Jurassic (Morrison Formation) of N Am
Characters:
Comments: A very primitive paracryptodire. Joyce 2007's computational-cladistic study was the first to assess the placement of Dorsetochelys delairi within a global testuidine analysis, revealing its placement as the sister taxon to Paracryptodira, although likely ancestral to the condition seen in more derived pancryptodires. It's basal position is also recovered by Sterli 2010, although the more detailed paracryptodire cladograms presented in Lyson & Joyce 2011 and Lyson et al 2011 presented Dorsetochelys as a basal Pleurosternid. This shows that advanced paracryptodire features evolved phyloparallelistically
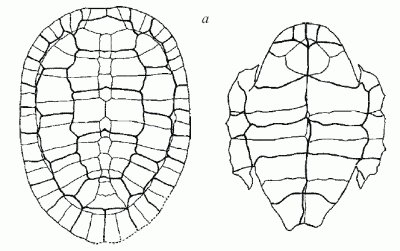
Pleurosternon bullocki (Owen, 1842) (= Pleurosternum ovatum Owen, 1853) from the Purbeck Limestone Formation (Berriasian, Early Cretaceous) of Durlstone Bay, Dorset County, UK. Carapace 50 cm, plastron 44 cm long as preserved) (Hirayama et al, 2000, p.185) |
Pleurosternidae Cope, 1868 : Compsemys, Glyptops, Pleurosternon
Range: Jurassic (Kimmeridgian) to Paleocene (Danian), Laurasia
Phylogeny: Paracryptodira : Compsemys + ((Arundelemys + Baenidae) + (Dorsetochelys + * : Pleurosternon + Glyptops))
Characters: $ Long basisphenoid reaching palatines (Gaffney, 1979a).
Comments: a possibly paraphyletic assemblage of Marine and Freshwater forms. Best known by Pleurosternon from the Early Cretaceous of England and Glyptops from the Late Jurassic of North America. Compsemys from the Late Cretaceous and Paleocene of North America might also be a member of this assemblage. Glyptops is sometimes placed in a separate family. The skull of Pleurosternon associated with postcranial fragments was originally referred to as Mesochelys.
Links: Glyptops - Wikipedia.
References: Gaffney, 1975. Hirayama et al, 2000 p.185
 |
| Carapace of Pleurosternon ovatum (syn of Pleurosternon bullocki) from the Purbeck Limestone, Swanage, Dorset. Original url, Joseph Dinkel, public domain, via Wikipedia |
Pleurosternon Owen 1853
Synonyms: Pleruosternum, Megasternum
Range: Late Jurassic (Tithonian) of England
Phylogeny: Pleurosternidae : Glyptops + *
Description: "Carapace much depressed, rounded posteriorly, without vacuities, and firmly united by a long bridge on each side with the plastron, which is also continuous. Surface of shell very finely pitted and deeply impressed with the suture-lines of the epidermal shields. Neural bones moderately elongated, hexagonal, with their anterolateral surfaces short. Mesoplastrals well developed, meeting in the middle line ; entoplastral relatively large ; xiphiplastrals deeply notched. No nuchal shield ; intergular shield not divided ; inframarginals present. Pectoral arch resembling that of the existing Chelys." (Zittel, Eastman, et al (1902), p.207)
Comments: From Wikipedia: - Pleurosternon fossils were first described by Richard Owen in 1841 under the living genus Platemys (Owen 1841). It was not until 1853 however, that it was published under the name Pleurosternon in a paper Owen presented to the Palaeontographical Society (Owen 1853)
Pleurosternon has a very depressed carapace, much flatter than similar genera, such as the North American Late Jurassic and Early Cretaceous Glyptops. Adults show little or none of the nuchal emargination that is more visible in juveniles. The Xiphiplastras also have a large, V-shaped notch near the back of the bone (Milner 2004). Together with Platychelys, it is one of the few turtle genera to exhibit the characteristics of both modern turtle suborders, the Cryptodira and the Pleurodira. (Hay 1908 p.45, Neaverson, 1955 p.114)
In Europe, most species of Pleurosternon are best known from southeast England's Purbeck Group and Portland stone, some specimens were even recovered in the Purbeck's type locality. (Neaverson, 1955 p.446) Several areas within the formation became noted by some for producing pleurosternon fossils. Among them were Swanage, Durlston Bay, Langton Matravers, and Herston. (Milner 2004 p.1448).
The genus has also been found in several rock formations from the upper Jurassic in both France and Spain (Pérez García et al 2009), and from the Cretaceous Wealden group of both England and northern Germany (Abel 1919 p.412). The Purbeck formation, at the time was a coastal region with a complex system of shallow lagoons that slowly lost their salinity over time (Radley date p 48). Similarly, the younger Wealden group was also coastal plain dominated by lagoons but with the addition of braided streams (Jackson 2008). The Portland stone, however is a maritime deposit of similar age near the Purbeck, most bones found there are interpreted as having washed out to sea from the Wealden or the Purbeck. Because of this, Pleurosternon has been described as a "shore-turtle". (Boule & Priviteau 1935)
Three species: Pleurosternon bullocki (Owen, 1853) (Type species), P. portlandicum (Lydekker, 1889), P. provinciale (Matheron, 1869)
Glyptops Marsh, 1890
Phylogeny: Pleurosternidae : Pleurosternon + *
Range: Late Jurassic to Early Cretaceous of North America
Comments: Adapted from Wikipedia: The generic name Glyptops (Greek for "grooved face") has been used to include a variety of fossil cryptodires from the Late Jurassic and Early Cretaceous of North America. Fossils have been found in South Dakota, Wyoming, Colorado, Utah, New Mexico, Oklahoma, and Texas from both the Morrison and Cedar Mountain formations. The type species is G. plicatulus, which was previously named Compsemys plicatulus by Edward Drinker Cope. Additional species have also been assigned to this genus: G. caelatus Hay, 1908, G. ornatus Marsh et al., 1988 (illustrated right), G. pervicax Hay, 1908, and G. utahensis Gilmore, 1916, although apart from G. plicatulus, no cladistic analysis has been done so it is not known how closely they are related or if some of them are synonyms. Marsh assigned this genus a new family Glyptopsidae but this ranking is unnecessary as it is unanimously placed in cladistic analyses with Pleurosternon (Joyce 2007, Lyson & Joyce 2009, Sterli 2010, etc) (although a mongeneric subfamily Glyptopsinae Marsh, 1890 would be fine). MAK130311
Arundelemys
Range: Mid Cretaceous (Albanian) to Late Eocene (Bartonian) of North America
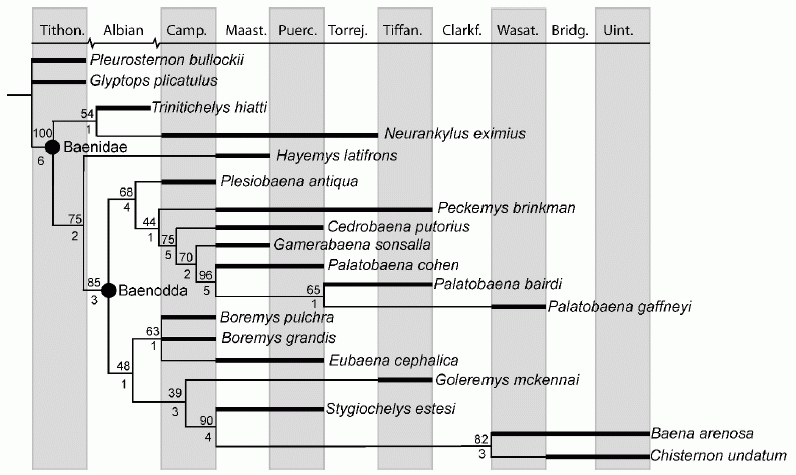
Baenidae Cope, 1882
Range: Mid Cretaceous (Albanian) to late-Mid Eocene (Uintian/Bartonian) of North America
Characters: absence of epipterygoids; acquisition of a contact between the first thoracic rib and the axillary buttresses; reduction on in width of the vertebrals, with vertebral scales 2 to 4 as narrow as, or narrower than, pleurals (Joyce 2007 p.65, Anquetin 2009, p.196)
Comments: A relatively conservative group of freshwater turtles common in Western Laurasia (North America) (Carroll 1988 p.212). Thanks to its prominent American fossil record, this is a cladistically well-studied group (e.g. Gaffney, 1972, Gaffney and Meylan 1988, Brinkman and Nicholls 1993, Lyson & Joyce 2009). As the chrono-cladogram above shows, most taxa ranged happily across the K-T boundary, unaffected by the end Cretaceous mass-extinction (Lyson & Joyce 2009, Lyson et al 2011). "Mainly found in deposits formed by river channels. They had slightly domed and spindle shaped shells, a very long tail and recurved claws at their feet, all suggesting that these were strong, bottom-walking turtles. Baenid turtles were characterized by a curious growth pattern: When adult size was reached, the individual bony plates forming the shell fused and the shell stopped expanding. After this, growth was invested in the thickening of the shell, especially of the lower plastron, resulting in an exceptional thickness of the lower shell in some specimens. Most baenids had massive jaws, and they probably fed on mollusks." (Jehle, 2006). The neck may have been slightly retractile.
Trinitichelys hiatti
Range: Albian of NA
Phylogeny: Baenidae : (Neurankylus + Baenodda) + *
Comments: The earliest and most basal Baenid(Lyson et al 2011), similar to Neurankylus .
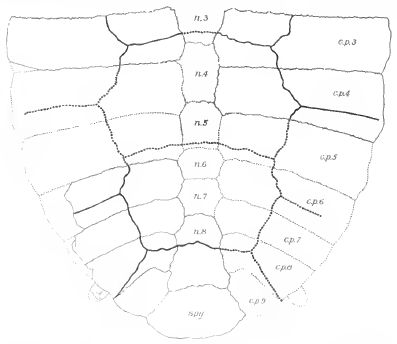 |
| Neurankylus eximius Hinder portion of carapace of type. From Lambe's figure. c.p.3, c.p.4, etc, costal plates; n.3, n.4, etc, neural plates; spy, suprapygal. From Hay 1908 p.94 |
Neurankylus eximius Lambe, 1902
Range: Late Cretaceous (Middle Campanian) to Mid Paleocene of North America
Phylogeny: Baenidae : Trinitichelys (Baenodda+ * )
Comments: Type specimen a partial carapace collected by Lawrence M. Lambe in 1901, from Judith River deposits (Middle Campanian) of the Red Deer River, Alberta. These seem to have come from an animal wioth a total carapce length of about 30 cm (Hay 1908 p.94). Lyson & Joyce (2009)'s cladogram has this species continuing to the mid Paleocene As species tend to average only several million years in range, it is not clear whether such a long-lived taxon belongs to a single species; perhaps there were distinctions that did not fossilise, e.g. in the genome or soft tissues. Or this may simply have bene a very succesful "living fossil". regardless, Together with Trinitichelys, Neurankylus represents the "Neurankylus lineage", one of the three primary Baenid evolutionary lineages, the other two being the Plesiobaena+Palatobaena and the Eubaena+Stygiochelys lineages (or subfamilies?). Shells of the Neurankylus lineage are large, with a finely crinkled sculpture pattern, and a complete ring of marginals, these being the basal condition for crown Testudines (Lyson & Joyce 2009). Later cladistic analyses shows this to be a paraphyletic, or in other words ancestral group, with the earlier (Albian, mid Cretaceous ) Trinitichelys being more basal (Lyson et al 2011), a nice example of stratigraphic congruency; although earlier analyses such as Joyce 2007 and Sterli 2010 recover Neurankylus as more basal, indicating the usual mosaic evolution and parallophyly. MAK130406
Baenodda Gaffney & Meylan, 1988
Range: Late Cretaceous (Campanian) to late-Mid Eocene (Bartonian) of North America
Phylogeny: Baenidae : Trinitichelys + (Neurankylus + * :Plesiobaena-Palatobaena lineage + Eubaena-Stygiochelys lineage)
Characters: absence or near absence of exposure of the prefrontal on the skull roof (Anquetin 2009, p.196)
Comments: The "higher" baenids. They marked an important ecological transition. Whereas basal forms like Arundelemys and Trinitichelys, each only known from a single specimen, were both found in claystone, indicative of a slow moving water environment. Neurankylus eximius appears in both claystones and mudstones, while baenodds are almost exclusively found in sandstones, indicating riverine environments. The paleoecological transition within Paracryptodira from low energy ponded environments of the earlier and more primitive forms to higher energy stream environments therefore took place near the Baenodda node (Lyson & Joyce 2011 p.798). During the latest Cretaceous this adaptive shift represented a classic evolutionary radiation attained a high level of diversity in the Maastrichtian, with nine taxa currently recognized (Lyson et al 2011). There are two main clades. A few stragglers continued through to the end of middle Eocene.
Plesiobaena-Palatobaena lineage
Range: Late Cretaceous (Campanian) to Early Eocene of North America
Phylogeny: Baenodda : Eubaena-Stygiochelys lineage + * : Plesiobaena + (Peckemys + (Cedrobaena + Palatobaena))
Characters: lack of supramarginal scutes, lack of a nuchal scute (scute located between the cervical scute and first vertebral scute), weak scalloping, anal scute located entirely on the xiphiplastron and reduced or absent gular scutes. - Lyson & Joyce 2009
Comments: of Maastrichtian and Paleocene taxa only Peckemys brinkman and Palatobaena cohen are known from shells. These are nearly identical except for the strong nuchal projection in the latter. It is assumed the other four taxa had similar shells, so identification of solitary shells beyond the lineage level is problematic. - Lyson & Joyce 2009
 |
| Plesiobaena antiqua Fig. 37. Carapace of holotype, 1/3 life size, n.1, etc., neural bones; nu.p. nuchal bone. Fig. 38. Anterior lobe of plastron. 1/2 life size, Both from drawings by Lambe.From Hay 1908 p.62 |
Plesiobaena antiqua (Lambe, 1902)
Range: Campanian to Paleocene of North America
Phylogeny: Plesiobaena-Palatobaena lineage : (Peckemys + (Cedrobaena + Palatobaena)) + *
Comments: Earliest and most primitive representative of the Plesiobaena-Palatobaena lineage. The type species, P. antiqua Lambe, 1902 is recorded from the Belly River Formation, Alberta (Lambe 1902), this being of Campanian age. An undescribed Maastrichian, and a named Paleocene species (P. putorius), extending the genus across the K-T boundary. A cladistic redescription by Lyson & Joyce 2009 reassigned these species to the new genera Peckemys and Cedrobaena respectively. This is in keeping with cladistic systematics which tends to appoint monospecific genera, as there is probably little doubt that were these extant taxa they would all be placed in the same genus. Obviously all such categorisation is arbitrary.
Peckemys brinkman Lyson & Joyce 2009
Range: Latest Maastrichtian to Late Paleocene of North America; distribution Hell Creek Formation (latest Maastrichtian) of North Dakota and Montana, Lance Formation (latest Maastrichtian) of Wyoming, Hannah Formation (Tiffanian - Late Paleocene) of Wyoming, and Laramie Formation (latest Maastrichtian) of Colorado. - Lyson & Joyce 2009
Phylogeny: Plesiobaena-Palatobaena lineage : Plesiobaena + (Cedrobaena + Palatobaena) + * )
Comments: identified as Plesiobaena-Palatobaena lineage based on features of the skull
Cedorbaena putorius
Range: Latest Maastrichtian to Late Paleocene of North America; distribution Hell Creek Formation (latest Maastrichtian) of South Dakota and North Dakota and Fort Union Formation (Torrejonian and Tiffanian) of Wyoming
Phylogeny: Plesiobaena-Palatobaena lineage : Plesiobaena + (Peckemys + (Palatobaena + * ))
Comments: Skull wedge shaped, larger than either Pl. antiqua or Pe. brinkman; similar in size to Palatobaena, Chisternon, and Stygiochelys.
Palatobaena
Range: Maastrichtian to Early Eocene of North America
Phylogeny: Plesiobaena-Palatobaena lineage : Plesiobaena + (Peckemys + (Cedrobaena + * ))
Comments: the most derived species of the lineage, represented by three presumably anagenetic species in stratigraphic and morphological sequence: P. cohen, P. bairdi, and P. gaffneyi. Co-existence with more primitivce species implies a number of co-existing lineages from a rapid mid/late maastrichtian evolutionary radiation. MAK130406
Eubaena-Stygiochelys lineage
Range: Late Cretaceous (Campanian) to late-Mid Eocene (Bartonian) of North America
Characters: (Boremys pulchra + Baena arenosa + Chisternon undatum) acquisition of supernumerary cervical scutes, an opisthocoelous caudal column, and the complete loss of cleithra, although the last characteristic is probably diagnostic for a more inclusive clade of baenids. - Joyce 2007 p.65; for Eubaena cephalica and Stygiochelys estesi) anterior supramarginal scales present, broken cervical scute, nuchal scute present, strongly scalloped posterior edge of shell, anal scute laps onto the hypoplastron, and well developed gular scutes - Lyson & Joyce 2009
Phylogeny: Baenodda : Plesiobaena-Palatobaena lineage + * : Boremys + (Stygiochelys + (Baena + Chisternon))
Boremys
Range: Late Campanian to Earliest Paleocene (Dinosaur Park, Judith River, Kirtland, Hell Creek and Fort Union formations) of N Am
Phylogeny: Eubaena-Stygiochelys lineage : Boremys + (Stygiochelys + (Baena + Chisternon)) + *
Comments: Includes the earliest representatives of the Eubaena-Stygiochelys lineage. The Maastrichtian Stygiochelys is a related form. Although B. pulchra and B. grandis have long been known from the Late Campanian. But recent discoveries show that Boremys continued into the late Maastrichtian and Early Puercian. The surprising failure to previously recognise this distinctive riverine genus in the Hell Creek Formation, which thanks to over a century of collecting provides one of the best understood terrestrial fossil vertebrate faunas, may be due to Boremys having a relatively thin shell in relation to other baenids, which in post-Campanian forms probably disarticulated more easily than those of other contemporaneous baenids. (Lyson et al 2011). The Maastrichtian Eubaena cephalica is a closely related species (Lyson & Joyce 2009)
Stygiochelys Gaffney & Hiatt, 1971
Range: Late Maastrichtian of Montana
Phylogeny: Eubaena-Stygiochelys lineage : Boremys + (Stygiochelys + (Baena + Chisternon)) + *
Comments: Presumably, the name "Turtle of the River Styx" refers to this riverine turtle being found in the Hell Creek formation, This animal had a sehell about 30 cm in length.
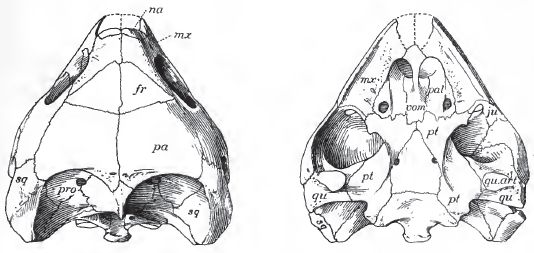 |
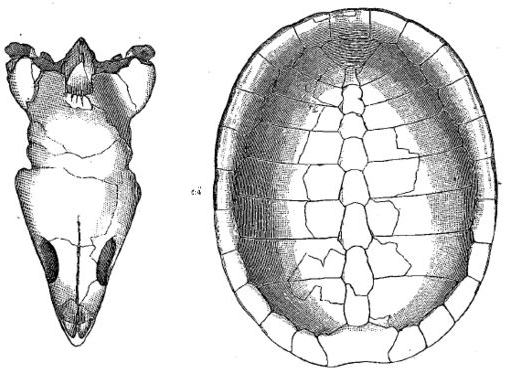
| Chisternon undatum, skull, 3/4 natural size, Bridger badlands (middle Eocene) of Wyoming. Left, top view, fr, frontal; mx maxilla; na, nasal; pa parietal; pro, prootic; sq, squamosal. Right, bottom view ju, jugal ; mx, maxilla ; pal, palatine; pt, pterygoid; qu, quadrate; qu. art, articulation of lower jaw; sq, squamosal; vom, vomer. From Hay 1908 p.89 (as Chisternon hehraicum - junior synonym) No. 5961 of the AMNH. |
"Baenini"
Range: Eocene of N Am
Phylogeny: Eubaena-Stygiochelys lineage : Boremys + (Stygiochelys + * : Baena + Chisternon)
Characters: reacquisition of a parietal squamosal contact; loss of splenials - Joyce 2007 p.65
Comments: This clade of Eocene paracryptodires (here informally given the rank of tribe) constitute the last representatives of the group. Like the contemporary Champsosaurs, they were Mesozoic survivors in a mid Paleogene world. MAK130407
|
|
| Baena arenosa left, Carapace, showing bones and scute areas. Right, Plastron, abj abdominal scute; an, anal scute; ent. entroplastroa; epi, epiplastron; fm, femoral scute; g, gular scute; hum^ humeral scute; hyo, hyoplastron; hypo, hypoplastron; infm, inframarginal scutes; mes, mesoplastron; pec, pectoral scute; xiph, xiphiplastron. Length 26 cm From Hay 1908 p.77 (as Baena riparia - junior synonym) |
Baena arenosa Leidy
Range: Early to Middle Eocene of N Am
Phylogeny: Eubaena-Stygiochelys lineage : Boremys + (Stygiochelys + (Baena + Chisternon)) + *
Description: "Intergular shield divided, and xiphiplastral with depressions which appear to have received the pubis and ischium. Mesoplastrals meeting at the middle line, but with the median ends much narrower than the outer ends." (Zittel, Eastman, et al (1902), p.207)
Comments: Originally included Cretaceous specuies, these have since been assigned to new genera.
Chisternon undatum Leidy
Range: Middle Eocene of North America
Phylogeny: Eubaena-Stygiochelys lineage : Boremys + (Stygiochelys + (Baena + Chisternon)) + *
Comments: Best known from the Middle Eocene Bridger Beds of Wyoming
checked ATW030512, last modified MAK100917
Page last modified MAK110922. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners

{kind=link}
{kind=link}