 We have discussed the pectoral limb of Eusthenopteron in some detail in connection with that genus. There is yet more that needs to be said, and we will say it. But first, a few words about the oft-forgotten pelvic limb.
We have discussed the pectoral limb of Eusthenopteron in some detail in connection with that genus. There is yet more that needs to be said, and we will say it. But first, a few words about the oft-forgotten pelvic limb.| Sarcopterygii | ||
| The Vertebrates | Elpistostegalia |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Teleostomi
├─Neopterygii
└─Sarcopterygii
├─┬─Onychodontiformes
│ └─Actinistia
└─Rhipidistia
├─Dipnomorpha
└─Tetrapodomorpha
├─Rhizodontiformes
└─Osteolepiformes
├─Tristichopteridae
└─Elpistostegalia
├─┬─Panderichthys
│ └─Elpistostege
└─┬─Livoniana
└─┬─Elginerpetonidae
└─┬─┬─Metaxygnathus
│ └─Ventastega
└─Tetrapoda
|
Contents
Overview |
We have discussed the pectoral limb of Eusthenopteron in some detail in connection with that genus. There is yet more that needs to be said, and we will say it. But first, a few words about the oft-forgotten pelvic limb.
As Andrews & Westoll (1970) point out, there is a strong tendency in the literature to treat the pelvic limbs -- if at all -- as simply the same as the pectoral limbs, only less so. This is perhaps understandable. Even in Panderichthys, the legs are very poorly developed compared to the arms, and good fossils of rhipidistian legs are even rarer than good fore-limbs. However, there are two issues that should be mentioned about legs: why are they so similar to arms, and why are they so different?
The similarities can be addressed relatively easily. Why develop an entirely new genetic program for hindlimbs? If something is worth doing, evolution ensures that it will be done twice, whether or not it is worth doing twice. Reproduction is the process of creating serial homologues of whole animals, and it shouldn't surprise us that the process also occurs on a smaller scale. With that said, Williston's "Law" often tends to reduce the number and similarity of serial homologues, so we might need to look further for an explanation of the continuing similarity of limbs. One factor may be the hypothesis of rhipidistian locomotion we have discussed elsewhere. If that hypothesis bears some relationship to reality, we would expect legs would remain similar to, but less developed than, arms because they were similar in function but lagged in evolutionary development.
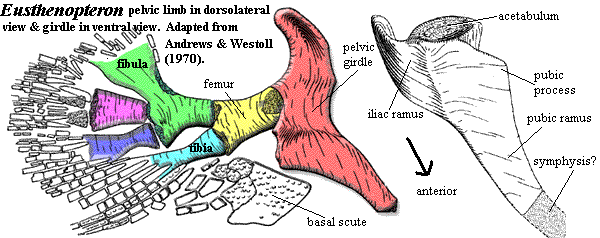
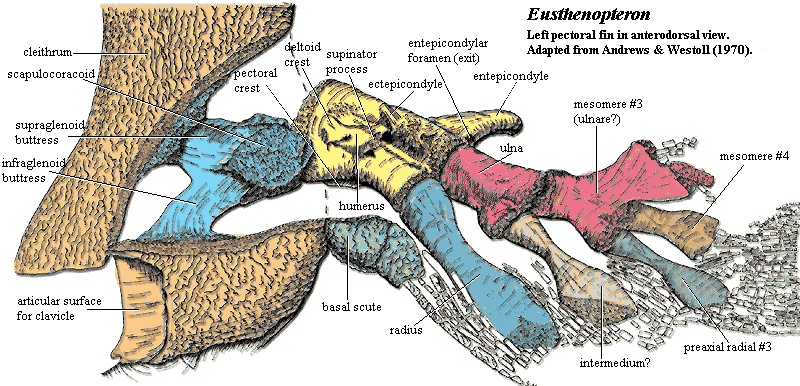
 The differences are perhaps more interesting. Compare the image of the hind limb of Eusthenopteron with the image of its forelimb. Note the peculiar shape of the one-piece pelvic girdle. Note also that the postaxial process is on the second mesomere, rather than the first and third, and the striking similarity of the fibula to the second mesomere in the fore limb. Many workers have noted the weird resemblance between the pelvic girdle and the humerus. Whether this is homology or coincidence is impossible to say at this point.
The differences are perhaps more interesting. Compare the image of the hind limb of Eusthenopteron with the image of its forelimb. Note the peculiar shape of the one-piece pelvic girdle. Note also that the postaxial process is on the second mesomere, rather than the first and third, and the striking similarity of the fibula to the second mesomere in the fore limb. Many workers have noted the weird resemblance between the pelvic girdle and the humerus. Whether this is homology or coincidence is impossible to say at this point.
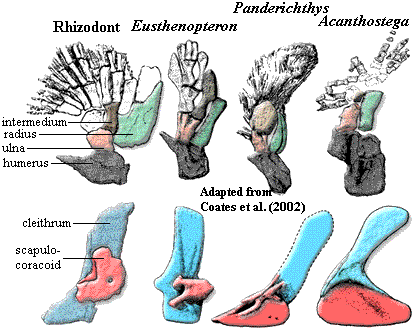
Returning to the pectoral limb, we can easily make out two general trends. First, the number of mesomere and radials is reduced. Coates et al. (2002). Second, and particularly in rhizodonts and elpistostegalians, we also observe an increase in bone surface ventral to the limb. In rhizodonts, the relevant bone is the ventral lamina of the cleithrum. In Panderichthys, it is the ventral lamina of the scapulocoracoid (the future coracoid plate). Both are probably associated with the development of massive ventral retractor/adductor musculature required by the increasing locomotor role of the pectoral fins. Janvier 1980). Development of the ectepicondyle and supinator processes suggests that the limb was also increasingly capable of more complex movements, particularly bending at the "elbow," [Andrews & Westoll (1970)] whether to react with substrate or (perhaps more likely) simply to create a more efficient recovery stroke. The limb also becomes longer and more ventral. These changes also have obvious relevance to the tetrapod-like locomotor style discussed previously. Finally, the scapulocoracoid consistently becomes more massive and the glenoid turns outward and/or downward. Coates et al. (2002). The range of motion permitted in the glenoid may also increase. All of these trends are explained by an evolutionary change in function from dorsoventral rotation, (associated with attitude control, steering and braking) to anteroposterior rotation (associated with locomotion. The pattern is somewhat mosaic, but seems to apply to the three principle tetrapodomorph lineages independently, suggesting an adaptive response. ATW031107.
GO TO PART 7. Conclusions, Scenarios, and Speculations
Most of our knowledge of the basal elpistostegalian fishes is due to Panderichthys, with some comparison possible to the closely-related Elpistostege and Livoniana, which is only known from the anterior jaw. Much of what we know of Panderichthys is due to the efforts of Prof. Emilia Vorobyeva (Эмилия Воробьева) and her co-workers, who have studied this genus intensively over almost 30 years. See, e.g., Vorobyeva 1980), Vorobyeva & Schultze (1991).
The skull of Panderichthys is strikingly close to the tetrapod model. The skull is low, with postfrontal ridges ("eyebrows") and dorsal orbits, suggesting the ability to see above the water line [LG04]. Many of the dermal bones articulate with interdigitating sutures. This is an important novelty of the Elpistostegalia, the implication being that the skull sometimes had to be supported against gravity [LG04]. The frontals are fully developed. The nasals are not yet consolidated, but are easily identified as a series of 2-3 bones lateral to a reduced field of small rostrals. The dermal intracranial joint is completely obliterated, although (as discussed previously) the internal separation of the braincase persists in non-tetrapod forms. The anterior margin of the parietals lies between the orbits. The spiracular (otic) notch is deep and forms a well-defined break between skull table and cheek. Primitively, the operculogular series is complete, but the operculars are distinctly narrowed. The internal structure of the teeth is essentially the same as in tetrapods. The jaw incrementally approaches the tetrapod condition -- which is a good thing, since most elpistostegalians are known only from jaw material.
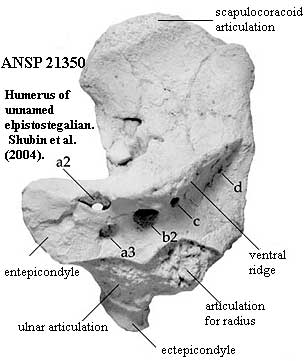 The scales lack cosmine and have developed a style of ornamentation closer to the tetrapod norm. The dorsal and anal fins are confluent with the caudal fin. In essence, only the paired fins and the tail fin remain. The hind limbs have begun to develop as major elements. In fact, the pelvic fins seem to lead the pectoral fins in evolutionary development [C+03]. The forelimbs are elongate and are positioned ventrally. In more derived forms, the limbs project laterally, and the range of motion of the humerus and femur are restricted. Importantly, the limbs are dorsoventrally flattened; and there are only three mesomeres, each quite specialized. [LG04]. This suggests that the fin musculature has already specialized into a variety of distinct components -- which, in turn would require major rewiring of the nervous system in this region. That impression is confirmed by the further development of various distal lumps, bumps and ridges on the stylopods. See image from [S+04]. These attest to the existence of distinct, large muscles which independently control the attitude of the zeugopod.
The scales lack cosmine and have developed a style of ornamentation closer to the tetrapod norm. The dorsal and anal fins are confluent with the caudal fin. In essence, only the paired fins and the tail fin remain. The hind limbs have begun to develop as major elements. In fact, the pelvic fins seem to lead the pectoral fins in evolutionary development [C+03]. The forelimbs are elongate and are positioned ventrally. In more derived forms, the limbs project laterally, and the range of motion of the humerus and femur are restricted. Importantly, the limbs are dorsoventrally flattened; and there are only three mesomeres, each quite specialized. [LG04]. This suggests that the fin musculature has already specialized into a variety of distinct components -- which, in turn would require major rewiring of the nervous system in this region. That impression is confirmed by the further development of various distal lumps, bumps and ridges on the stylopods. See image from [S+04]. These attest to the existence of distinct, large muscles which independently control the attitude of the zeugopod.
Some workers apparently regard the Elpistostegalia as a monophyletic group of Baltic fishes similar to Panderichthys. See brief discussion in [R+03]. This seems to be an unsettled issue, and given the recent indications that stem and early tetrapods were both diverse and common in the latest Devonian, it may be unsafe to say anything much about phylogenetic details. Mind you, this will not confine our usual rash speculations in the slightest. However, it ought to. And so, having both acknowledged and renounced our moral obligations in the matter, we will move on to the details. ATW031107, revised ATW050414.
 Elpistostegalia: = Panderichthyidae = Panderichthyida)
Elpistostegalia: = Panderichthyidae = Panderichthyida)
Range: from the latest Middle Devonian (late Givetian) [C97]
Osteolepiformes:::: Tristichopteridae + *: (Panderichthys + Elpistostege) + (Livoniana + Elginerpetonidae + ((Metaxygnathus + Ventastega) + Tetrapoda))).
Characters: head enlarged, flattened & spade-shaped [AM94] [CA96]; preorbital skull lengthened [AM94]; snout dermal bones primitively complex, with median rostral, anterior, median and posterior postrostrals, 3 paired nasals, with 1st nasal fused to premaxilla [SA85]; maxilla posterodorsal process absent [CA96$]; primitively, nares surrounded by lateral rostral & anterior tectal [SA85]; nares very close to margin of jaw [VS91] [CA96$] [C96$]; skull roof bones with tetrapod-like pattern [SA85]; skull roof flat [VS91]; distinct paired frontals [SA85] [AM94] [CA96$]; frontals present, flanked by elongate prefrontals (homologous to supraorbital 1) [SA85]; prefrontal does not reach nares [SA85]; frontals also flanked by elongate postfrontals (homologous to supraorbital 2) [SA85]; frontals closely sutured to parietals between orbits with dermal intracranial division absent [VS91]; postfrontal reaches further anteriorly than parietals [SA85]; distinctive "eyebrows" formed by postfrontals [SA85] [CA96]; postfrontal forms dorsal margin of orbit [SA85]; orbits dorsal & closely spaced, with flat or concave interorbital skull [VS91] [AM94] [CA96$]; orbits with concave interorbital region [AM94]; jugal meets quadratojugal & excludes squamosal from maxilla [VS91] [CA96$]; distinct intertemporals, supratemporals, & tabulars present [SA85]; postorbital may reach spiracular notch [SA85]; extratemporal 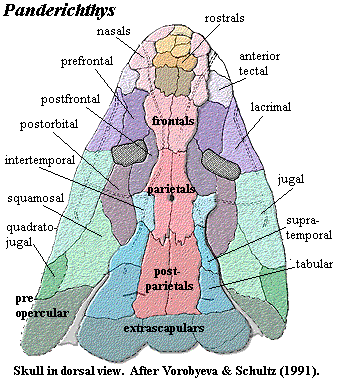 and postspiracular plate absent [SA85]; large spiracular notch extends posteriorly to form squamosal embayment "otic notch") [VS91] [CA96$]; complete operculogular set present [A+00]; basicranial fenestra present [A+00]; intracranial joint present [A+96]; intracranial joint immobilized externally [AM94] [CA96$] [A+00]; lateral ethmoid does not contact palatoquadrate [SA85]; medial ethmoid contact with palatoquadrate synchondrosal [SA85]; external intracranial joint lost [VS91] [CA96$]; basicranial fenestra & arcual plates absent [CA96$]; nasal capsules primitively well-ossified [SA85]; nasal capsules small & well separated [VS91]; vomers with posterior processes meeting at midline & excluding pterygoids from midline [SA85] [CA96]; Meckelian bone exposed dorsal to prearticular [A+00]; primitively, large median gular present [SA85] [CA96$]; dentition labyrinthodont; teeth with deep infolding at base & internally polyplocodont [SA85]; anterior tusks in lower jaw [SA85]; pleurocentra not ossified [CA96$]; ribs expanded? questioned by [C02]; anal and dorsal fins absent [VS91] [AM94] [CA96$] [C96$]; fin basal scutes absent [R+03]; epichordal lepidotrichia more developed than hypochordal [CA96$]; large scapulocoracoid plate [C96$]; scapulocoracoid having single attachment to cleithrum [SA85]; supraglenoid and infraglenoid buttresses present [C96]; humerus somewhat elongated; scales rhombic, with coarse, bony ornamentation [SA85]; humerus with strap-like head [S+04]; humerus with anterior keel(??) [C96$]; ectepicondyle reaches distal end of humerus [R+03]; dermal bones & scales without cosmine [SA85].
and postspiracular plate absent [SA85]; large spiracular notch extends posteriorly to form squamosal embayment "otic notch") [VS91] [CA96$]; complete operculogular set present [A+00]; basicranial fenestra present [A+00]; intracranial joint present [A+96]; intracranial joint immobilized externally [AM94] [CA96$] [A+00]; lateral ethmoid does not contact palatoquadrate [SA85]; medial ethmoid contact with palatoquadrate synchondrosal [SA85]; external intracranial joint lost [VS91] [CA96$]; basicranial fenestra & arcual plates absent [CA96$]; nasal capsules primitively well-ossified [SA85]; nasal capsules small & well separated [VS91]; vomers with posterior processes meeting at midline & excluding pterygoids from midline [SA85] [CA96]; Meckelian bone exposed dorsal to prearticular [A+00]; primitively, large median gular present [SA85] [CA96$]; dentition labyrinthodont; teeth with deep infolding at base & internally polyplocodont [SA85]; anterior tusks in lower jaw [SA85]; pleurocentra not ossified [CA96$]; ribs expanded? questioned by [C02]; anal and dorsal fins absent [VS91] [AM94] [CA96$] [C96$]; fin basal scutes absent [R+03]; epichordal lepidotrichia more developed than hypochordal [CA96$]; large scapulocoracoid plate [C96$]; scapulocoracoid having single attachment to cleithrum [SA85]; supraglenoid and infraglenoid buttresses present [C96]; humerus somewhat elongated; scales rhombic, with coarse, bony ornamentation [SA85]; humerus with strap-like head [S+04]; humerus with anterior keel(??) [C96$]; ectepicondyle reaches distal end of humerus [R+03]; dermal bones & scales without cosmine [SA85].
Notes: [1] Since [SA85] believed that Panderichthyidae included only Panderichthys and Elpistostege, we have adapted their characters slightly to make them more compatible with the current belief that the group includes Tetrapoda.
Image: Panderichthys adapted from 2001, who adapted it from [J96].
References: Ahlberg & Milner (1994) [AM94]; Ahlberg et al. (1996) [A+96], Ahlberg et al. (2000) [A+00], Clack (1997) [C97]; Clack (2002) [C02], Cloutier & Ahlberg (1996) [CA96], Coates 1996) [C96]; Coates et al. 2002) [C+03], Janvier (1996) [J96]; Long & Gordon (2004) [LG04]; Ruta et al. (2003) [R+03]; Schultze & Arsenault (1985) [SA85], Shubin et al. (2004) [S+04]; Vorobyeva & Schultze (1991) [VS91]. ATW050413
checked ATW051025
{kind=link}