Sarcopterygii: Actinistia: Diplocercides
Taxa on This Page
- Diplocercides X
Frankly, the purpose of this section is to showcase an image from Jarvik 1980) and to talk a bit about the jaw articulation it illustrates so well. Jarvik's images are some of the finest anatomical drawings we have run into in some time. Jarvik's style of labeling was remarkably unattractive. Worse, we have, as usual, meddled with color and textures, in addition to playing with labels in our never-ending struggle against incomprehensible anatomical abbreviations. Even so, Jarvik's illustrations of the skull of "Nesides" (= Diplocercides) are so wonderfully clear that they withstand even these sustained attempts at uglification. Oddly, Jarvik may never have seen this particular skull. The specimen -- all 4 cm of it -- was ground to dust by Erik Stensiö, who used the grinding method to create an enlarged, detailed, wax model of the braincase. Stensiö's wax model was the actual source of the image.
This section is also our first effort written for a new server, a venue which gives us virtually unlimited space for images. We are sorely tempted to abuse this privilege, but there are countervailing considerations. Not the least of these is a nagging suspicion that some readers may, unaccountably, fail to share our boundless enthusiasm for the skull of Diplocercides. Nevertheless, it is a real pleasure to have enough detail, from enough different angles, that we can see how the whole thing fits together. Those of us who don't happen to have Paleozoic fish skulls around the house often have a hard time visualizing how the typical anatomical illustration translates into a working organism. Accordingly, we have compromised by providing supplemental images of the neurocranium in dorsal and ventral views. In addition, see occipital view of neurocranium at Actinistia.
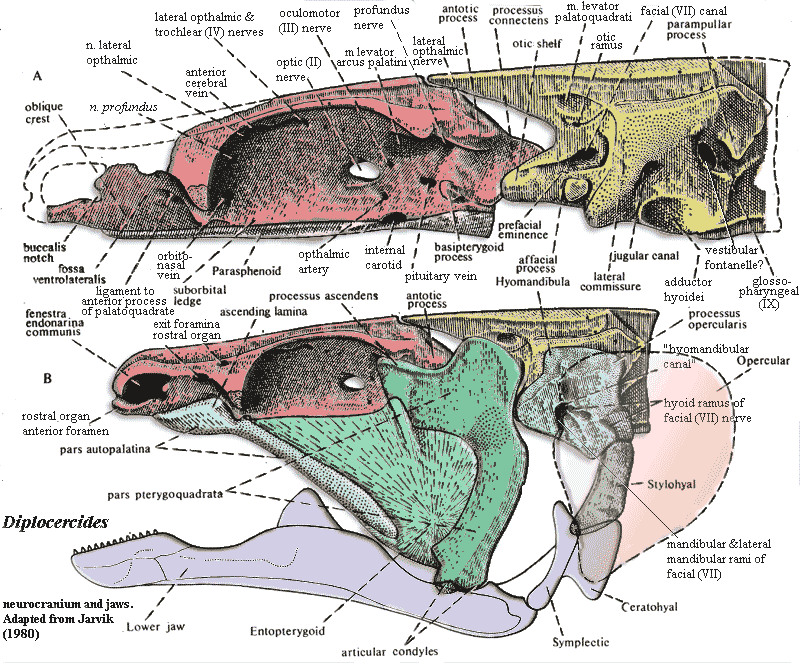
So, having digested this massive graphic, what does it mean? Actually a good bit. Working from front to back, we will take in just a few of the sights -- mostly relating to the upper jaw articulation:
1. Actinistians have a unique sensory organ. It is called the rostral organ for the excellent reason that it is found in the rostrum. Internally, it looks like a third nasal capsule, squeezed in between, and on top of, the other two. It opens anteriorly by a pair of foramina which look like a third pair of nares, located in front of the anterior nares. However, unlike a nasal capsule, the rostral organ also has external exits -- or, at least, it has two pairs of additional openings. Jarvik 1980). The organ is filled with a gelatinous substance in Latimeria, and presumably nothing macroscopic actually exits the organ. It is said that the organ "may serve an electrosensory function" -- which is what ichthyologists always say when they have no idea what function a given piece of anatomy actually performs.
2. Actinistians have no maxilla. The general belief is that the maxilla was lost at the inception of the actinistian line. We question whether any actinistian ancestor ever had a maxilla homologous to the tetrapod maxilla. Consider the "maxilla" of Psarolepis. Is that really a maxilla? Might it be better regarded as a jugal or quadratojugal? Or might it be a neomorph, serially homologous to an opercular series? More than one of the above? At any rate, it is unsafe to assume that the actinistians "lost" any of the dermal skull bones which osteolepiforms/tetrapods have, but coelacanths lack. The points of departure for inference ought to be Psarolepis andStyloichthys which probably has no maxilla of any sort). Conclusions about the plesiomorphic state based on Holoptychius, the classical model, need serious revision based on the discoveries of the last decade.
3. At any rate, absent a maxilla, the palatoquadrate is forced to make do by clinging onto the neurocranium directly at various points. Anteriorly, actinistians develop a strong ligamentous connection between palate and braincase, as do many other fishes. See, e.g. Huysentruyt & Adiaens (2005) (Corydoras, a catfish); Motta & Wilga (1999) (Ginglymostoma, nurse shark). This anterior portion of the palatoquadrate is known as the autopalatine. Jarvik, incidentally, regarded the autopalatine as the remnant of a premandibular arch. It is still possible -- if rarely -- to find this segmentalist speculation repeated in papers published a quarter century later. Huysentruyt & Adiaens (2005). This is a subject too large for the present context. Still, it is interesting that, after going thoroughly out of style for some time, this proposition may turn out to have been partially correct, if in a very unexpected way. At any rate, the autopalatine is often found as a separate ossification in actinopterygians, where it is closely associated with the palatine.
 4. Note that the upper jaws are essentially parallel, as is the pattern in all surviving groups of gnathostomes. This feature can be best seen in the supplemental figures. Parallel upper jaws are clearly not the ancestral condition. In Psarolepis and Styloichthys the sphenethmoid portion of the braincase expands widely, forward of the intracranial joint. The border of the sphenethmoid region is marked off by a pair of sphenethmoid buttresses which form a V-shaped outline in ventral view, diverging anteriorly. As a consequence, if the orbits were placed in their usual (for living gnathostomes) position, the eyes would be very widely separated and would tend to face posterolaterally. This, presumably, is why basal gnathostomes possessed eyestalks and/or anteriorly placed orbits. Among actinistians, the very early Miguashaia uniquely retains a short, wide snout with orbits placed anteriorly (i.e., anterior to the widest point of the skull). However, this is enough for us to conclude that Miguashaia had the primitive type of braincase, and that actinistians probably evolved parallel jaws independently of other fishes -- by parallel evolution, so to speak.
4. Note that the upper jaws are essentially parallel, as is the pattern in all surviving groups of gnathostomes. This feature can be best seen in the supplemental figures. Parallel upper jaws are clearly not the ancestral condition. In Psarolepis and Styloichthys the sphenethmoid portion of the braincase expands widely, forward of the intracranial joint. The border of the sphenethmoid region is marked off by a pair of sphenethmoid buttresses which form a V-shaped outline in ventral view, diverging anteriorly. As a consequence, if the orbits were placed in their usual (for living gnathostomes) position, the eyes would be very widely separated and would tend to face posterolaterally. This, presumably, is why basal gnathostomes possessed eyestalks and/or anteriorly placed orbits. Among actinistians, the very early Miguashaia uniquely retains a short, wide snout with orbits placed anteriorly (i.e., anterior to the widest point of the skull). However, this is enough for us to conclude that Miguashaia had the primitive type of braincase, and that actinistians probably evolved parallel jaws independently of other fishes -- by parallel evolution, so to speak.
5. The middle part of the upper jaw is not actually part of the palatoquadrate. It is a dermal bone known as the entopterygoid, or just plain pterygoid. That is, it is either homologous to the dermal bone which dominates the tetrapod palate, or a really good imitation. See the glossary entry at pterygoid for a long-winded explanation.
6. The anterior process of the autopalatine is sometimes, as in the image, referred to as an "ascending lamina." This is unfortunate, since one of the trademark characters of the Sarcopterygii (perhaps, but not necessarily, a basal synapomorphy) is the ascending process of the palatoquadrate, which isn't the same thing at all. The ascending process of the palatoquadrate is a feature of the metapterygoid, the upper part of the pterygoquadrate. It is wonderfully displayed in Diplocercides as a multiple articulation (again, see the glossary entry at pterygoid for the definitions of these terms). The ascending process fits into a slot in the braincase posterodorsal to the orbit, while the supratemporal process (also called suprapterygoid process) and the antotic process of the sphenethmoid articulate in a palatoquadrate depression just posterior to the ascending process. (The suprapterygoid process is not labeled in the figure, but is the process just anterior to the antotic process, separated from the latter by a strong notch.) The palatoquadrate then rises dorsally again to receive the m. levator palatoquadrati which originates on the otoccipital portion of the braincase. This conformation seems to be what Schultze (1973) suggests that he found in Grossius.
7. In Diplocercides, the metapterygoid also retains the basicranial articulation through the basipterygoid process. This is actually a little unusual for actinistians. Latimeria and Mesozoic coelacanths generally lose the basipterygoid process, so that the principle motion is around the dorsal articulation. The mechanics of the actinistian jaw are poorly understood (Forey, 1998). However, it may make better mechanical sense to place the axis of rotation dorsally, since this gives the levator palatoquadrati more mechanical advantage (but also a slower response).
8. Finally, note that there is no direct connection between the hyomandibular and the jaw. Instead, the short, blocky hyomandibula connects with the lower jaw independently, through a chain of bones articulating on a separate, retroarticular bone. Actinistians, uniquely, ossify the Meckelian cartilage at three points: the mentomeckelian near the lower jaw symphysis, the articular where the upper jaw connects through a double articulation, and the retroarticular which accommodates the symplectic connection with the hyomandibula. Experimental post-mortem manipulation of the jaw causes the intracranial joint to flex; and some theories of actinistian jaw mechanics make much of this. However, it seems likely that actinistians have more than one jaw closing mechanism and considerable flexibility in combining the various mechanical possibilities. ATW051223.
 Diplocercides: Stensiö 1922). D. ("Holoptychius") keysari van Koenen, 1895 (= Nesides schmidti); D. davisi Moy-Thomas, 1937; D. ("Nesides?") heiligenstockiensis Jessen, 1966; D. jaeckeli Stensiö, 1922.
Diplocercides: Stensiö 1922). D. ("Holoptychius") keysari van Koenen, 1895 (= Nesides schmidti); D. davisi Moy-Thomas, 1937; D. ("Nesides?") heiligenstockiensis Jessen, 1966; D. jaeckeli Stensiö, 1922.
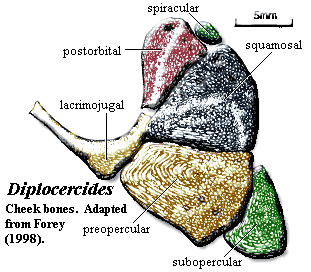
Characters: head generally low [F98]; cranial ornament of tubercles & ridges generally longitudinal [F98]; lateral rostral is simple tube around infraorbital canal [F98]; antorbital unremarkable [F98]; skull table bones with simple abutting sutures except between parietals (overlap) [F98]; skull table flat [F98]; skull roof ornamented with dense covering of tubercles [F98]; four large median internasals, flanked by 4 pairs. of nasals, flanked by 4 pairs. of tectals [F98]; two pairs. nasals [F98]; parietal descending process absent [LJ88]; about 6 supraorbitals [F98]; supraorbital canal straddles sutures between tectal and parietal series [F98]; supraorbital canal pore openings large [J73]; intracranial border straight [F98]; postparietal unusually flat [F98]; supratemporal and postparietal descending laminae absent (plesiomorphic) [LJ88] [F98]; postparietal and supratemporal form straight posterior border [F98]; 3 extrascapulars with large laterals [F98]; Cheek: cheek region tall and anteroposteriorly short [J73]; orbit large [J73]; orbit with numerous sclerotic ossicles [F98]; cheek bones as in figure, covering entire cheek and tightly sutured [F98]; cheek bones completely ornamented with small tubercles & ridges [F98]; lacrimojugal with distinct ventral "elbow," does not extend significantly onto rostrum [F98]; squamosal triangular, overlapping postorbital, & reaching skull roof [F98]; squamosal tall posteriorly [J73]; squamosal triangular with peak facing anteroventrally [J73]; spiracular tiny & scale-like [F98]; quadratojugal absent [F98] (contra [J73] quadratojugal present as partially separated posterior margin of preopercular); preopercular overlaps
 subopercular [F98]; Braincase: see also discussion above; braincase completely ossified except interorbital & ethmoid areas largely perichondral [F98]; opisthotic with vertical crest bearing postotic process for epaxial trunk muscle attachment [F98]; saccular chamber of otic capsule large, pierced by large fenestra which may be homologous to vestibular fontanelle [F98]; lateral commisure with jugular canal as in osteolepiforms [F98]; afacial eminence bearing spiracular tooth plate [F98]; otic shelf extends anteriorly into process articulating with processus connectens [F98] (see image at Actinistia); other features as indicated in braincase image above;
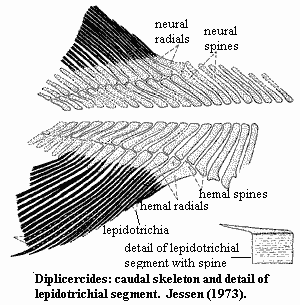
3 postorbital processes: (a) antotic (b) basipterygoid & (c) suprapterygoid process (just anterior to antotic, separated by notch) [F98]; antotic process large [LJ88]; basipterygoid process present, well-developed, lying directly ventral to antotic process [F81] [F98]; oculomotor (III) nerve enters orbit directly from interorbital region [LJ88]; nasal capsule with 3 foramina to orbit: (a) superficial ophthalmic branch of dorsal lateral line nerve (b) profundus nerve & (c) orbitonasal artery & vein [F98]; nasal capsule with fossa ventrolateralis for autopalatine, containing ridge for ligaments to autopalatine [F98]; Palate: parasphenoid very long, broad, extending below nasal capsules [F98]; pterygoid with fold or process mid-way along dorsal margin [F+00]; metapterygoid with deep socket for articulation with the basipterygoid process [F+00]; Lower jaw: mentomeckelian unusually long [F98]; dentary relatively long, occupying 1/3 of jaw length, without hooked process or dentary pore [F98]; dentary heavily ornamented laterally [F98]; 2-3 normal coronoids with simple tooth plates [F98]; principal coronoid relatively low with small coronoid expansion [F98]; splenial markedly shorter than dentary [F98]; prearticular shallow [F+00]; articular & retroarticular fused [F98] [F+00]; retroarticular deep [F+00]; retroarticular without distinct cartilage-capped articulation for symplectic [F+00]; ornament on angular, dentary & splenial of elongate ridges, as on gular [F98]; Dentition: tiny conical teeth in rows on anterior coronoids, dorsal pterygoid, & ventral surangular [F98]; parasphenoid with small villiform teeth for most of length [F98]; teeth ankylosed to dentary [F+00]; Axial: neural & hemal arches & spines very broad distally and may abut one another [J73] [F98]; lepidotrichia of unpaired fins segmented, sometimes bifurcate in middle and distal sections, especially in anal fin [J73]; 2nd dorsal and anal fins directly opposite each other [J73]; hemal spines long and numerous just anterior to caudal fin [J73]; caudal fin symmetrical & tri-lobed [F+00]; first radials of upper & lower caudal lobe rest on two spines [J73]; caudal fin with rays/radials ~2:1 [J73] [F98]; "caudal lepidotrichia in which successive segments interlock by means of step-like sutures" (D.
heiligenstockiensis) [F+00] [J73] [1]; Appendicular: pectoral girdle narrow [F+00] contra [F98]: pectoral girdle broad); clavicle, cleithrum & extracleithrum ornamented with ridges running length of bones [J73] [F98]; extracleithrum narrow & splint-like [F+00] (contra [F98]: extracleithrum unusually large) Other. scales ellipsoidal, with ridges running longitudinally [J73].
subopercular [F98]; Braincase: see also discussion above; braincase completely ossified except interorbital & ethmoid areas largely perichondral [F98]; opisthotic with vertical crest bearing postotic process for epaxial trunk muscle attachment [F98]; saccular chamber of otic capsule large, pierced by large fenestra which may be homologous to vestibular fontanelle [F98]; lateral commisure with jugular canal as in osteolepiforms [F98]; afacial eminence bearing spiracular tooth plate [F98]; otic shelf extends anteriorly into process articulating with processus connectens [F98] (see image at Actinistia); other features as indicated in braincase image above;
3 postorbital processes: (a) antotic (b) basipterygoid & (c) suprapterygoid process (just anterior to antotic, separated by notch) [F98]; antotic process large [LJ88]; basipterygoid process present, well-developed, lying directly ventral to antotic process [F81] [F98]; oculomotor (III) nerve enters orbit directly from interorbital region [LJ88]; nasal capsule with 3 foramina to orbit: (a) superficial ophthalmic branch of dorsal lateral line nerve (b) profundus nerve & (c) orbitonasal artery & vein [F98]; nasal capsule with fossa ventrolateralis for autopalatine, containing ridge for ligaments to autopalatine [F98]; Palate: parasphenoid very long, broad, extending below nasal capsules [F98]; pterygoid with fold or process mid-way along dorsal margin [F+00]; metapterygoid with deep socket for articulation with the basipterygoid process [F+00]; Lower jaw: mentomeckelian unusually long [F98]; dentary relatively long, occupying 1/3 of jaw length, without hooked process or dentary pore [F98]; dentary heavily ornamented laterally [F98]; 2-3 normal coronoids with simple tooth plates [F98]; principal coronoid relatively low with small coronoid expansion [F98]; splenial markedly shorter than dentary [F98]; prearticular shallow [F+00]; articular & retroarticular fused [F98] [F+00]; retroarticular deep [F+00]; retroarticular without distinct cartilage-capped articulation for symplectic [F+00]; ornament on angular, dentary & splenial of elongate ridges, as on gular [F98]; Dentition: tiny conical teeth in rows on anterior coronoids, dorsal pterygoid, & ventral surangular [F98]; parasphenoid with small villiform teeth for most of length [F98]; teeth ankylosed to dentary [F+00]; Axial: neural & hemal arches & spines very broad distally and may abut one another [J73] [F98]; lepidotrichia of unpaired fins segmented, sometimes bifurcate in middle and distal sections, especially in anal fin [J73]; 2nd dorsal and anal fins directly opposite each other [J73]; hemal spines long and numerous just anterior to caudal fin [J73]; caudal fin symmetrical & tri-lobed [F+00]; first radials of upper & lower caudal lobe rest on two spines [J73]; caudal fin with rays/radials ~2:1 [J73] [F98]; "caudal lepidotrichia in which successive segments interlock by means of step-like sutures" (D.
heiligenstockiensis) [F+00] [J73] [1]; Appendicular: pectoral girdle narrow [F+00] contra [F98]: pectoral girdle broad); clavicle, cleithrum & extracleithrum ornamented with ridges running length of bones [J73] [F98]; extracleithrum narrow & splint-like [F+00] (contra [F98]: extracleithrum unusually large) Other. scales ellipsoidal, with ridges running longitudinally [J73].
Links:
References.
Notes: [1] from [J73], with customary apologies for our worthless translation of the German: "Most segments of the anterior lepidotrichia in the lower caudal lobe appear to bear an enamel-covered spine. These spines emerge in each case from the external enamel-covered surface of the segments on the margins of both caudal lobes and are oriented so as to overlie distally the following segment."
References: Forey (1981) [F81]; Forey (1998) [F98]; Forey et al. (2000) [F+00]; Jessen 1973) [J73]; Lelièvre & Janvier (1988) [LJ88]. ATW060116.
{kind=link}