
Holoptychius. Artwork by Nobu Tamura, 2009. Wikipedia, GNU Free Documentation/Creative Commons Attribution license.
| Sarcopterygii | ||
| The Vertebrates | Allenypterus |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Teleostomi
├─Neopterygii
└─Sarcopterygii
├─┬─Onychodontiformes
│ └─Actinistia
│ ├─Diplocercides
│ └─┬─Hadronector
│ └─┬─┬─Allenypterus
│ │ └─Holopterygius
│ └─┬─Lochmocercus
│ └─┬─Polyosteorhynchus
│ └─┬─Rhabdoderma
│ └─Coelacanthiformes
└─Rhipidistia
├─Dipnomorpha
└─┬─Rhizodontiformes
└─Osteolepiformes
├─Tristichopteridae
└─Elpistostegalia
├─Panderichthys
└─Tetrapoda
|
Contents
Overview |
Holoptychius. Artwork by Nobu Tamura, 2009. Wikipedia, GNU Free Documentation/Creative Commons Attribution license. |
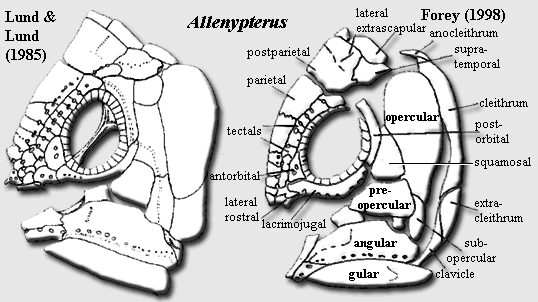
Allenypterus: Melton, 1969. A. montanus Melton, 1969.
Range: Mississippian (Serpukhovian) of North America (Montana).
Phylogeny: Actinistia :::::: Holopterygius + *.
Characters: 3-15 cm [LL84]; skull bones except premaxilla heavily ornamented with broad, flat-topped ridges [LL84] [F98]; skull deeper than long [F98]; median skull table bones all anamestic [LL84]; premaxilla narrow & splint-like [LL84] [F98]; small median rostral present [LL84] (questioned by [F98]); star-shaped rostrals [F98]; premaxilla without perforations or emargination for pores [LL84]; parietonasal shield 3X longer than postparietal [F98] [5]; 1-2 pr nasals & 2 pr. parietals [F98]; parietals with interdigitating suture [F98]; tectal series with at least 7 elements [F98] supraorbital bones may fuse with medial skull table elements [LL84]; supraorbital sensory line with pores along tectal/parietal sutures [F98]; dermal intracranial joint straight [F98]; postparietals almost square, strongly vaulted [F98]; supratemporal unreduced [LL84]; "posttemporal is in contact with the posterolaterally expanded tabular" [LL84] [2] (contra [F98]: supratemporal & no others lateral to postparietals); uncertain if spiracular present [F98] (noting disagreement); 3 (or 5?) extrascapulars present, scale-like & weakly associated with postparietal [F98]; probably no descending processes on postparietal or supratemporal [F98]; lateral rostral small, unornamented and bearing small ventral process [F98]; antorbital small, triangular, wedged between lacrimojugal & a tectal, bearing two foramina [F98]; lacrimojugal tubular [F98];  Cheek: orbit large, with numerous sclerotic ossicles [F98]; cheek bones thin, broadly overlapping [LL84] (contra [F98]: only overlap is squamosal, pre- & subopercular overlap onto opercular); cheek bones unornamented [F98]; squamosal triangular, relatively small [F98]; subopercular present [LL84]; subopercular small, scale-like & overlapped by preopercular [F98] [6]; Lower Jaw: jaw deep [F98]; dentary unornamented, without hook-shaped process [F98]; dentary dorsally concave, rod-like, much longer than splenial [F98]; splenial very small [F98]; principal coronoid relatively small, with rounded apex [F98]; angular does not reach symphysis [F98]; angular lateral surface ornamented with prominent ridges tending to run longitudinally [F98]; splenial short, deep & ornamented [F98]; jaw articulation at level of posterior orbital margin [F98] [4]; articular-retroarticular fused & very large [F98]; premaxilla with row of fine teeth [LL84] [F98]; Axial: body shape tall, compressed, dacriform [LL84] [F98] [1]; notochord arched high over abdomen [F98]; 1st dorsal with long base plate [F98]; 1st dorsal fin rays "short, well-articulated and of uniform height" [LL84] [3]; 2nd dorsal small, located in anterior half of body [F98]; 2nd dorsal fin articulates directly above base plate (compare Caridosuctor) [LL84]; anal fin small [F98]; anal fin supported by 1st hemal spine [LL84]; caudal with greatly extended epicaudal and vestigial hypocaudal [LL84]; upper caudal lobe begins in anterior half of body as low fringe of fin rays [F98]; caudal fin ray/radial ratio >1:1 [LL84] [F98]; caudal terminal lobe lepidotrichia absent [LL84] (contra [F98]: a few rays); lower caudal lobe restricted to a few posterior fin rays but with numerous (40) radials [F98]; pectoral fin high, about halfway up body wall; median ventral row of 6-9 thick scales on ventral surface between clavicles and pelvic girdle [LL85] [F98]; pelvic girdles are tall & triangular, do not articulate at midline [LL84]; scales small, ornamented with many longitudinal ridges which may converge posteriorly [F98].
Cheek: orbit large, with numerous sclerotic ossicles [F98]; cheek bones thin, broadly overlapping [LL84] (contra [F98]: only overlap is squamosal, pre- & subopercular overlap onto opercular); cheek bones unornamented [F98]; squamosal triangular, relatively small [F98]; subopercular present [LL84]; subopercular small, scale-like & overlapped by preopercular [F98] [6]; Lower Jaw: jaw deep [F98]; dentary unornamented, without hook-shaped process [F98]; dentary dorsally concave, rod-like, much longer than splenial [F98]; splenial very small [F98]; principal coronoid relatively small, with rounded apex [F98]; angular does not reach symphysis [F98]; angular lateral surface ornamented with prominent ridges tending to run longitudinally [F98]; splenial short, deep & ornamented [F98]; jaw articulation at level of posterior orbital margin [F98] [4]; articular-retroarticular fused & very large [F98]; premaxilla with row of fine teeth [LL84] [F98]; Axial: body shape tall, compressed, dacriform [LL84] [F98] [1]; notochord arched high over abdomen [F98]; 1st dorsal with long base plate [F98]; 1st dorsal fin rays "short, well-articulated and of uniform height" [LL84] [3]; 2nd dorsal small, located in anterior half of body [F98]; 2nd dorsal fin articulates directly above base plate (compare Caridosuctor) [LL84]; anal fin small [F98]; anal fin supported by 1st hemal spine [LL84]; caudal with greatly extended epicaudal and vestigial hypocaudal [LL84]; upper caudal lobe begins in anterior half of body as low fringe of fin rays [F98]; caudal fin ray/radial ratio >1:1 [LL84] [F98]; caudal terminal lobe lepidotrichia absent [LL84] (contra [F98]: a few rays); lower caudal lobe restricted to a few posterior fin rays but with numerous (40) radials [F98]; pectoral fin high, about halfway up body wall; median ventral row of 6-9 thick scales on ventral surface between clavicles and pelvic girdle [LL85] [F98]; pelvic girdles are tall & triangular, do not articulate at midline [LL84]; scales small, ornamented with many longitudinal ridges which may converge posteriorly [F98].
Links: Allenypterus montanus, IIDB - "Living Fossils"; IIDB - View Single Post - The Coelacanth.
Notes: [1] or perhaps not. From all appearances, Allenypterus was, like many Bear Creek actinistians, ventrally broad & flat. Thus, it might be more accurate to say that Allenypterus was "semidacriform," that is, it looked like a tear drop cut in two. [2] We're not sure what to make of this, since our understanding is that actinistians are not known to have either of these bones. [3] This is inconsistent with the figure in [LL84], which shows anteriorly long lepidotrichia, becoming short posteriorly. [4] Elsewhere in the same work, Forey indicates that the jaw articulation is "behind the level of the operculum" which may be a more accurate description. However, given the distorted shape of the head, use of the usual anteroposterior landmarks for "level" easily leads to ambiguous references. [5] Elsewhere in the same work, Forey states that the "parietal" [meaning postparietal?] shield is 45% of the length of the parietonasal shield, making the ratio 2:1. [6] Elsewhere in the same work, Forey states that the preopercular is "a very large triangular element."
References: Forey 1998) [F98]; Lund & Lund (1984) [LL84]; Long (1995) [L95].. ATW060116.
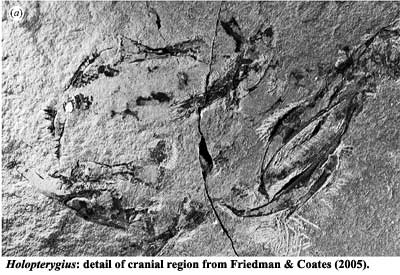 Holopterygius: a twisted Fish
Holopterygius: a twisted FishWe have no real quarrel with the reconstruction of Holopterygius given by Friedman & Coates (2005) -- and certainly no quarrel we'd have a chance of winning. However, it needs to be pointed out that the only known specimen of this fish is extremely scrappy and hard to interpret. It was originally described by Jessen (1973) as "Pices: incertae sedis" -- possibly a tarsiid actinopterygian. To make matters worse, the only specimen of Holopterygius is clearly the worse for wear after 35 years. A close comparison of Jessen's photograph of the cranial area with the same area in Friedman & Coates shows marked deterioration and loss of material.
Holopterygius is a sort of cubist coelacanth. The difficulty of saying much of anything in particular about the skull is obvious from the figure. The dentition and shape of the jaw are generally coelacanth-like, but the length of the tooth-bearing region of the mandible is suspicious. The dentary makes up less than half the length of the jaw in all other actinistians. We attempted to reconstruct the large piece in the upper central part of the image as an actinistian braincase. The result was almost, but not quite, convincing. Like most of the rest of Holopterygius, it gives the general impression of being an actinistian, but resists unambiguous assignment of its component parts.
 As Friedman & Coates point out, the main lesson of Holopterygius is that Paleozoic coelacanths were plainly a diverse lot, with a much wider range of body forms than we might expect from looking only at Mesozoic specimens. The Mesozoic forms (with the possible exception of Libys) all look more or less like the extant Latimeria. Friedman & Coates attempt to quantify this morphological diversity using a "relative warps analysis." This involves mapping the bodies of interest on a common coordinate plane, normalized for size, and noting the coordinates of selected landmarks. A software package then attempts to determine a series of functions which warp the coordinate plane so that the observed variation is explained with the minimum number of warps. As one might expect, the mathematics are similar to principal components analysis, so that the first warp is the single function which can explain the greatest amount of the morphological variance. The second warp is the single warp function which explains the greatest amount of the remaining variance, and so on.
As Friedman & Coates point out, the main lesson of Holopterygius is that Paleozoic coelacanths were plainly a diverse lot, with a much wider range of body forms than we might expect from looking only at Mesozoic specimens. The Mesozoic forms (with the possible exception of Libys) all look more or less like the extant Latimeria. Friedman & Coates attempt to quantify this morphological diversity using a "relative warps analysis." This involves mapping the bodies of interest on a common coordinate plane, normalized for size, and noting the coordinates of selected landmarks. A software package then attempts to determine a series of functions which warp the coordinate plane so that the observed variation is explained with the minimum number of warps. As one might expect, the mathematics are similar to principal components analysis, so that the first warp is the single function which can explain the greatest amount of the morphological variance. The second warp is the single warp function which explains the greatest amount of the remaining variance, and so on.
This is a relatively undemanding application of quantitative morphometric tools. Friedman & Coates are simply looking for indicators of morphological diversity. However, the technique clearly has much broader potential application. See, e.g., Tang & Pantel (2005), Anderson & Roopnarine (2005). Accordingly, we dug a bit deeper. Frieß 2003) includes a brief explanation (mercifully light on statistical jargon) of the technique in a paleontological context, using the same software package used by Friedman & Coates. Frieß's treatment also raises a number of methodological issues worth consideration.
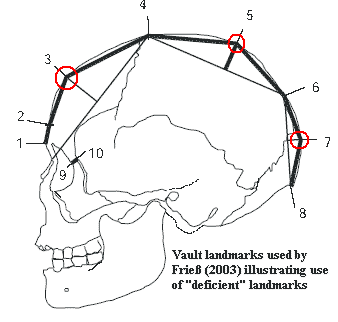 One issue of particular relevance to vertebrates is the form of the warp function. Apparently the model used in most paleontological applications is the "thin spline" -- a mathematical model which penalizes large changes between closely spaced landmarks. While morphological gradualism is a good bet for single bones, or unitary structures such as shells, we question whether it is appropriate for whole vertebrates. If two landmarks are, for example, at the opposite ends of adjacent long bones, a change in the angle of articulation may drastically alter the relative position of the two landmarks without necessarily causing any significant change in the overall morphology of the animal.
One issue of particular relevance to vertebrates is the form of the warp function. Apparently the model used in most paleontological applications is the "thin spline" -- a mathematical model which penalizes large changes between closely spaced landmarks. While morphological gradualism is a good bet for single bones, or unitary structures such as shells, we question whether it is appropriate for whole vertebrates. If two landmarks are, for example, at the opposite ends of adjacent long bones, a change in the angle of articulation may drastically alter the relative position of the two landmarks without necessarily causing any significant change in the overall morphology of the animal.
Another set of problems relate to size. First, what does it mean to normalize specimens for size? Equal surface area for flat specimens? What about volume, a critical, but rarely available, datum? Second, morphological parameters frequently exhibit some positive or negative allometry with size; and size varies with ontogeny as well as phylogeny. Most coelacanths are, mercifully, about the same size -- at least within an order of magnitude ~5-50 cm for mature specimens. However, Latimeria gets far larger, and may approach 200 cm. It is possible to apply a rough correction for this distortion Frieß, 2003: 65), but there is no indication whether such a correction was applied by Friedman & Coates.
Frieß was working with human skulls -- nearly a worst case for spherical distortion. We would not expect to see this type of error in the typical Bear Gulch coelacanth fossil, because it is entirely two-dimensional. However, this may be a significant source of error when three-dimensional material is available.
Finally, Frieß discusses the use of "deficient" landmarks, landmarks derived from the geometry of other landmarks. These are more easily illustrated than described. In the image, landmarks 3, 5 and 7 are "deficient" because they represent "points of maximum curvature" between two other landmarks, i.e., where the actual outline of the skull departs maximally from the chord drawn between the two reference landmarks. This type of landmark is deprecated in the theoretical literature. However, Frieß notes that they (a) make no difference to the clustering of his data points and (b) enhance the ability to describe the difference between clusters. The problem is somewhat analogous to the use of correlated characters in a typical parsimony-based cladistic study. In essence, the use of deficient characters "weights" portions of the anatomy because, for example, the information given by landmark 5 is partially redundant with the information given by landmarks 4 and 6. However, all landmarks have some degree of correlation, and the use of intermediate landmarks of this type may be used (as Frieß uses them) to compensate for the absence of clear anatomical markers in some regions of the skull. In other words, without the "deficient" landmarks, portions of the shape would be underweighted. This is clearly a judgment call. ATW060116.
 : Jessen, 1973. H. nudus Jessen, 1973.
: Jessen, 1973. H. nudus Jessen, 1973.
Range: Middle or Late Devonian (Givetian or Frasnian) of Europe Germany)
Phylogeny: Actinistia :::::: Allenypterus + *.
Characters: ~7 cm [J73]; skull bones "paper thin" and unornamented [J73]; opercular rectangular, taller than long [FC05]; large otoliths present [FC05]; parasphenoid splint-like with well-developed hypophyseal foramen [FC05]; triangular palatoquadrate [FC05]; ectopterygoid present, toothed [FC05]; mandible with long, slender "anterior ramus" (= dentary) [FC05]; mandible with large dorsally directed central projection (principal coronoid) [FC05]; mandible with posterior ramus (articular-retroarticular) [FC05]; marginal teeth simple & conical [J73]; well-developed premaxillary dentition with 4 large teeth [FC05]; body form eel-like [FC05]; body leaf-shaped, with greatly elongated caudal region [FC05]; vertebral centra unossified except possible parapophyses [FC05]; neural arches relatively wide, arranged in contact with each other, each with a flat, posterodorsally inclined spine. [J73]; hemal arches triangular, with longer, posteroventrally oriented spines posterior to the abdominal region [J73]; fin rays thin & thread-like, without segmentation or branching [J73]; no ossified ribs [FC05]; spines and neural/hemal arches co-ossified [FC05]; median fin line continuous from posterodorsal to shoulder girdle around the body to anal region [J73]; strongly asymmetrical caudal fin lobes (c.f. Miguashaia, Allenypterus), with well-developed ventral lobe [FC05]; dorsal fin extends far anterior (as Allenypterus) [FC05]; caudal fin with basal : radial 1:1 and rays: radials > 1:1 [FC05] (accord [J73]: ray: radial ratio about 2:1); anterodorsal radials stand quite erect, while more posteriorly radials are more posterodorsally oriented [J73] dorsal radials terminate bluntly, proximally and distally [J73]; pectoral girdle gracile (all actinistians except Miguashaia) [FC05]; Pectoral girdle posteriorly convex, high and rather narrow, showing fine external striations, more robust than skull bones [J73] [1]; anocleithrum unbranched [FC05]; extracleithrum present [FC05]; keel-like scales along ventral margin from skull to mid-abdominal region (c.f. Allenypterus) [FC05]; scales otherwise absent from body [J73].
Notes: [1] Jessen notes the similarity to actinistians in the shoulder girdle.
References: Friedman & Coates (2005) [FC05]; Jessen 1973) [J73]. ATW060126.
Lochmocercus: Lund & Lund, 1984. L. aciculodontus Lund & Lund, 1984. [1]
Range: Mississippian (Serpukhovian) of North America (Montana).
Phylogeny: Actinistia ::::: Polyosteorhynchus + (Caridosuctor + (Rhabdoderma + Coelacanthiformes))) + *.
Characters: skull roof bones sparsely ornamented with coarse tubercles, with other skull bones bearing more ornament [LL84]; median skull table bones all anamestic [LL84]; supraorbital bones may fuse with medial skull table elements [LL84]; supratemporal unreduced [LL84]; antorbital small [F98]; cheek bones abut closely [LL84]; cheek completely covered with tightly sutured bones [F98]; all cheek bones heavily ornamented [F98]; lacrimojugal not expanded anteriorly [F98]; anterior margin of postorbital approximates line of intracranial joint [LL84]; cheek bones closely fitted [F98]; subopercular present [LL84]; jaw shallow [F98]; oral margin of dentary continuous with coronoids (like Miguashaia?) [LL84]; dentary without hooked process [F98]; angular coronoid process well-developed [F98]; dentary and coronoids with large teeth [LL84] [F98] dentary teeth ankylosed to bone [F98]; fin rays with 2:1 ratio to bases [LL84] [F98]; caudal fin small & rounded [F98]; 2nd dorsal fin articulates directly above base plate (compare Caridosuctor) [LL84]; anal fin supported by 1st hemal spine [LL84]; anocleithrum present, stout & may have contacted skull [LL84]; extracleithrum large and strongly integrated [LL84]; pelvic plate quadrangular & lacks "significant" perichondral bone [LL84] pelvic limb elements ossified [LL84].
Notes: [1] Comment from [LL84: 243]: "L. aciculodontus is the rarest and most poorly preserved of the Bear Gulch coelacanths. Neither the rostral region nor the axial skeleton are completely preserved on any specimen. It is, however, the only coelacanth in the fauna with a 2/1 ratio of caudal fin rays to endoskeletal supports."
References: Forey 1998) [F98]; Lund & Lund (1984) [LL84]. ATW060119.
Polyosteorhynchus: Lund & Lund, 1984. P. simplex Lund & Lund, 1984.
Range: Mississippian (Serpukhovian) of North America (Montana).
Phylogeny: Actinistia ::::: Caridosuctor + (Rhabdoderma + Coelacanthiformes)) + *.
Characters: 3.5-19 cm, with maximum height anterior to 1st dorsal [LL84]; head nearly as deep as long [F98]; skull bones relatively thin, ornamented with coarse, low, flat-topped tubercles or ridges [LL84]; rostral bones sparsely ornamented [LL85]; dorsal margin of premaxilla emarginated for at least one pore [LL84]; rostral small and postrostrals may be absent [LL84] [1]; antorbital large [F98]; median skull table bones all anamestic [LL84]; median skull table elements not symmetrical [LL84]; no enlarged, clearly defined parietals [LL84] [2]; postparietal shield 2/3 length of parietonasal shield, flat, with straight posterior margin [F98]; spiracular present [F98]; supratemporal unreduced [LL84]; supratemporal (and possibly also postparietal) descending process(es) present [LL85]; 3 or 5 extrascapulars [F98]; cheek bones deeply overlapping [LL84]; cheek completely covered with large, tightly sutured, overlapping bones [LL85] [F98]; all cheek bones heavily ornamented [F98] (contra [LL85]: lightly ornamented); lacrimojugal not expanded anteriorly [F98]; sclerotic ring well-developed [F98]; postorbital extends over intracranial joint [LL84]; postorbital narrow [F98]; may be 2 squamosals in some individuals [LL84]; preopercular large, trapezoidal [F98]; squamosal & preopercular overlap opercular & subopercular [LL85]; opercular narrow, tall [LL85] [F98]; opercular conspicuously ornamented with concentric pattern of large tubercles [LL85]; subopercular present, tall and somewhat triangular [LL85]; opercular & subopercular overlap shoulder girdle [LL85]; small toothed palatine may be present [LL85]; metapterygoid wide & stout [LL85]; jaw shallow [F98]; dentary and splenial of same length [F98] dentary without hook-shaped process [F98]; articular & retroarticular fuse late in ontogeny [F98] or are separate [LL85]; premaxilla with 3-4 large teeth [LL84] [F98]; dentary with tiny, villiform teeth [F98]; dentary plate with small teeth [LL85]; prearticular with longitudinal rows of denticles [LL85]; 2nd dorsal fin articulates directly above base plate (compare Caridosuctor) [LL84]; anal fin supported by 1st hemal spine [LL84]; caudal fin outline approaches square [LL84] (contra [F98]: rounded); caudal ray/radial ratio about 1:1 [LL85]; anocleithrum with stout, spike-like vertical process [LL85]; dorsal part of cleithrum short & weakly ornamented with vertical ridges [LL85]; cleithrum with long anteroventral process articulating with dorsal arm of clavicle [LL85]; extracleithrum large & heavily ornamented [LL85]; clavicles expanded & loosely associated at ventral midline [LL85]; pelvic plates mid-abdominal, long & narrow, without posteromedial articular process [LL84]; ossified swim bladder present [LL85] [F98].
Links: Polyosteorhynchus. But, even here, we cannot locate a good image of this fish -- particularly the rather interesting pectoral girdle.
Notes: [1] because of the general uncertainties of nomenclature in Actinistia, we are not certain what bone is being referred to. We believe this is also what we would call the (median) rostral. If so, and if the lateral rostral (one of the "postrostrals") is actually absent, then the marginal pores on the premaxillae may represent the nares. [2] Apparently something functionally equivalent is formed by fusion of smaller bones in large specimens [LL85].
References: Forey 1998) [F98]; Lund & Lund (1984) [LL84], Lund & Lund (1985) [LL85]. ATW060119.
checked ATW041103