Sarcopterygii: Actinistia: Hadronector
Taxa on This Page
- Hadronector X
Descriptions
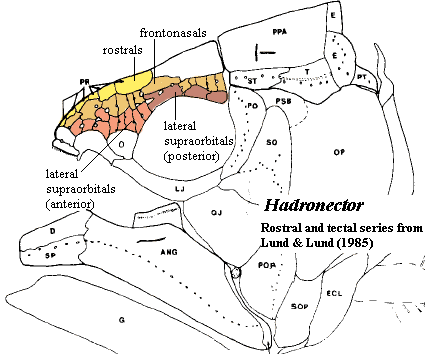 One of the more frustrating experiences which awaits the student of coelacanth paleontology is the impossibility of reconciling Lund & Lund (1985) with Forey (1998). This conflict comes to a head, so to speak, over the tectal/superorbital series. In general, the Lunds find that there are two marginal series of bones that parallel the median pairs of skull table bones of the nasoparietal shield. Forey usually finds a single series. The Lunds are wont to find additional dermal bones in other places, as well; but the nasoparietal shield is the location in which they have their most consistent differences with Forey.
One of the more frustrating experiences which awaits the student of coelacanth paleontology is the impossibility of reconciling Lund & Lund (1985) with Forey (1998). This conflict comes to a head, so to speak, over the tectal/superorbital series. In general, the Lunds find that there are two marginal series of bones that parallel the median pairs of skull table bones of the nasoparietal shield. Forey usually finds a single series. The Lunds are wont to find additional dermal bones in other places, as well; but the nasoparietal shield is the location in which they have their most consistent differences with Forey.
But why do we care whether these particular folks butt heads over tectals? Consider the protagonists. In this section we cover ten Paleozoic coelacanths -- almost all of the reasonably known actinistians of that Era. Of these ten, five are known only from Bear Gulch. Richard Lund is the Baron of Bear Gulch, a locality with a unique and poorly understood taphonomic style. On the one hand, Bear Gulch fishes were killed very quickly -- including the famous example of a pair of stethacanthid sharks fossilized in flagrante delicto. Preservation of gut contents and even pigments is fairly common. On the other hand, the specimens are completely flattened and, thus, have undergone a certain amount of physical distortion and crushing. Lund has probably been involved in the substantial majority of vertebrate finds at Bear Gulch and undoubtedly has a better handle on this peculiar mode of preservation than anyone else.
Peter Forey we have already introduced as the man who wrote The Book on fossil coelacanths. He publishes relatively little, but his papers are massively documented, carefully reasoned, and universally respected. The general feeling is that he is correct and that Lund, who is better known for work on chondrichthyans, may not have had a sufficient handle on the sarcopterygian material at Bear Gulch.
Due to some distressing childhood trauma, now suppressed, or perhaps some innate moral deformity, the right/wrong thing doesn't always resonate with us. When trained observers consistently differ in their observations, and almost always differ in the same directions, one often finds that there is an explanation other than chance or skill. Sometimes, that explanation is more interesting than a binary standard of right or wrong. In any case, we decided to have a close look at the evidence in a test case, the supraorbitals (tectals) of Hadronector.
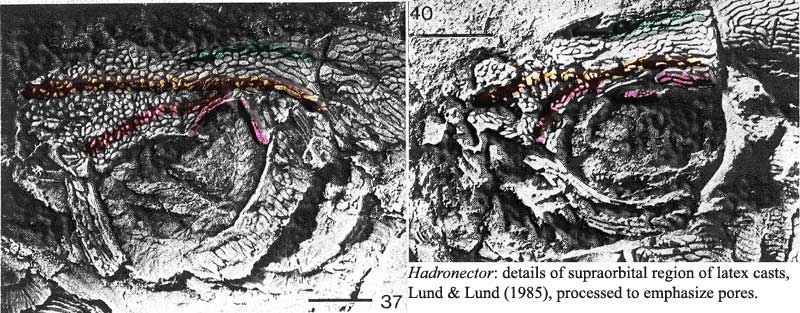
We picked Hadronector because it is strikingly unstriking, a Joe Average among coelacanths. In addition it is the only Bear Gulch coelacanth in which all specimens are essentially the same size. Presumably, they were spawned and grew up elsewhere. Lund & Lund (1985). This uniformity reduces the chance that differing observations might be due to variation in developmental stage among specimens. Our own bias was that Forey would probably turn out to be correct here, simply because Hadronector is superficially similar to garden-variety Mesozoic actinistians.
 Unfortunately, it not quite so easy to dismiss these observations. What appears to be a doubled anterior pore line is not difficult to make out. As advertised, the lower line terminates in two or three larger bones without pores which frame the orbit posterodorsally. Indeed, there is some indication of an additional line of some kind (faintly outlined in green) medially. A somewhat similar general pattern seems to be present in Allenypterus, in the color image
Unfortunately, it not quite so easy to dismiss these observations. What appears to be a doubled anterior pore line is not difficult to make out. As advertised, the lower line terminates in two or three larger bones without pores which frame the orbit posterodorsally. Indeed, there is some indication of an additional line of some kind (faintly outlined in green) medially. A somewhat similar general pattern seems to be present in Allenypterus, in the color image
On the other hand, Forey states: "The parietonasal series consists of small nasals, at least three, which lie anteriorly and two pairs of parietals which lie above the orbit. Lund and Lund 1985: fig. 44) restore a double series of bones lying lateral to the parietals. I find no evidence of this" [F98: 55]. Forey's statement that Hadronector has two pairs of parietals is inconsistent even with his own figure 3.3B, which shows only one pair. He reconstructs the parietals as being flanked only by a dotted line -- presumably indicated uncertainty that any tectals flanked the parietals, and with no sensory pores posterior to the mid-point of the orbit.
As both papers point out, Hadronector has heavy ornamentation on the skull roof, and it is impossible to tell fracture from suture. One is forced to work by proxies such as pore lines and by comparing lines of fracture/suture between specimens. We may also have chosen an inauspicious test case, since Forey had only limited material available to him [F98: 316]. Still, it would seem that Forey's phrase, "no evidence," is overstated.
However, let us return briefly to the implications of the "green line." This probably only reflects the median suture between the parietals. The implication is that a line of "pores" may actually represent a suture line. In some fossils, cylindrical matrix infilling structures allow one to spot sensory pores unambiguously, but these are generally absent in Hadronector [LL85: 28]. In that case -- and if we assume (1) that the red line is a bona fide sensory line contained entirely by single bones, but (2) the orange line is a suture line -- we are left with something very like Forey's reconstruction. The first assumption is not unreasonable for Paleozoic actinistians in which the sensory line pores are often large and surrounded by a single tectal, rather than being small, numerous and marginal.
We are thus left with considerable uncertainty. To make matters worse, the actinistian literature includes any number of references to cranial dermal bones being split or partially doubled or, alternatively, fused and of a dual nature. The whole discussion has a vaguely theological flavor, rather like the debates over the dual nature of Christ which led to the First Council of Nicea. Possibly this issue could be resolved in the same manner by Prelates of the Paleozoic at the next SVP meeting, after which all might enjoy a spectacle in which either Forey or Lund, like Arius the Libyan, would be declared Anathema and exiled to Illyria, all his writings being cast into the flames. ATW060128.
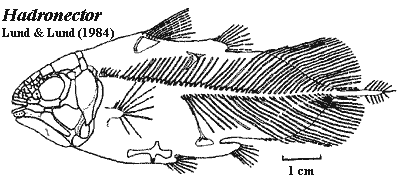 Hadronector:Lund & Lund, 1984. H. donbairdi Lund & Lund, 1984.
Hadronector:Lund & Lund, 1984. H. donbairdi Lund & Lund, 1984.
Range: Mississippian (Serpukhovian) of North America (Montana).
Phylogeny: Actinistia :::: (Allenypterus + Holopterygius) + (Lochmocercus + (Polyosteorhynchus + (Caridosuctor + (Rhabdoderma + Coelacanthiformes))))) + *.
Characters: 3.5-12 cm; head deep [F98]; all dermal skull bones heavily ornamented with coarse tubercles, closely spaced and tend to run into ridges posteriorly [LL84] [F98]; premaxilla with narrow dorsal lamina emarginated for anterior opening to rostral organ [LL84] [F98]; premaxillae do not meet on midline [LL85]; ?rostral series with 6 bones, extending to orbit [LL84]; parietonasal shield only slightly longer than postparietals [F98]; median skull table bones all anamestic [LL84]; at least 3 median internasals [F98] [3]; at least 3pr nasals & 2pr parietals [F98] [5]; three parallel tectal series [LL85] [1]; supraorbitals very narrow over orbits [LL84]; supraorbital bones may fuse with medial skull table elements [LL84]; spiracular present, small [LL85] [F98]; dermal intracranial joint straight [F98]; intertemporal unreduced [LL84] (contra [F98]: intertemporal absent) [4]; supratemporal with curved anterior margin [LL85]; posttemporal extends posterior to lateral extrascapulars [LL84]; 3 extrascapulars, with median element small forming straight posterior margin [F98]; lateral rostral with strong ventral process [F98]; antorbital large, with
2 posterior rostral organ foramina [F98]; cheek bones sutured or overlapping [LL84] [F98]; overlaps between at least postorbital, squamosal & preopercular [F98]; cheeks bones ornamented with flat tubercles running into ridges [F98]; lacrimojugal narrow, with small posterior expansion [F98]; postorbital narrow [F98]; postorbital anterior margin even with intracranial joint [LL85]; squamosal narrow and tall [F98] spiracular present, quite small [F98]; opercular & subopercular heavily ornamented [LL85]; subopercular present, much smaller than opercular [LL84] [LL85]; subopercular anamestic [F98]; quadratojugal absent [F98] (noting disagreement on this point); outer jaw bones heavily ornamented [F98]; palate with small, triangular autopalatine, tooth-bearing palatine & ectopterygoid [LL85]; pterygoid with concentric rings of fine denticles [LL85]; metapterygoid short & stout [LL85]; metapterygoid articulates with antotic process, with basipterygoid absent [LL85]; quadrate robust, strongly convex posteriorly [LL85]; lateral face of mandible strongly ornamented [LL85]; dentary same size as splenial, without hooked process or dentary pore [F98]; angular with low coronoid process [LL85] [F98]; angular dips about 40° downwards from plane of dentary & splenial [LL85]; premaxilla with tubercular ornament grading into fine marginal teeth [LL85] [2]; premaxilla with many small teeth [F98]; toothed coronoids? (i.e. marginal teeth, not tooth plates) [LL85] (this isn't clear); dentary with small marginal teeth [LL85]; almost toothless suction feeder; body short & stocky [LL84]; body scales ornamented as head scales, with ridges tending to converge posteriorly [LL85]; First 3 neural arches broadly in contact, only slight median crest [LL85]; neural arches 4-7 successively more arched and more widely spaced [LL85]; only 22 precaudal segments [LL85]; fin ray/radial ration 1:1 [LL85]; 1st & 2nd dorsal fins closely spaced, near mid-point of body length [F98]; 1st dorsal with long base plate [F98]; 2nd dorsal fin articulates directly above poorly ossified base plate (compare Caridosuctor) [LL84] [LL85]; anal fin with base plate supported by 1st "full-length" hemal spine [LL84] [LL85]; caudal fin outline can be almost square [LL84]; pectoral girdle elements ornamented with vertical ridges [LL85]; pectoral girdle located low on flank [LL85]; supratemporal present [LL85] [1]; anocleithrum with S-shaped dorsal process [LL85]; cleithrum long [LL85]; extracleithrum small, short [LL84] [LL85]; clavicle short [LL85]; pelvic plates located in mid-abdomen, anteriorly forked, broad [LL84]; pelvic plates with posteromedial articulating process [LL84] [LL85] body ornament of densely spaced ridges of roughly equal length [F98].
Links: Hadronector donbairdi Bear Gulch site)
Notes: [1] These are items which Forey either disputes or would presumably dispute, based on his general statements in [F98]. For additional discussion of the arrangement of tectals, see discussion above. [2] This also occurs in some extant African catfishes. [3] Possibly the same as the rostral series of [LL85]. [4] As discussed elsewhere, we follow Forey's nomenclature. Forey's supratemporal, i.e. the most posterior (and generally the only) dermal bone lateral to the postparietal, is called a tabular by Lund & Lund. The bones, if any, anterior to the supratemporal are called intertemporals by Forey and supratemporals by Lund & Lund. [5] As noted above, this is inconsistent with Forey's figure, which shows only one parietal.
References: Forey 1998) [F98]; Lund & Lund (1984) [LL84]; Lund & Lund (1985) [LL85]. ATW060116.