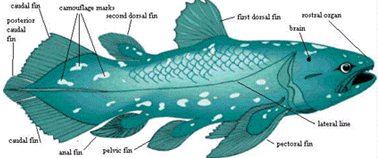
 Actinistia: (Coelacanthamorpha)
Actinistia: (Coelacanthamorpha)| Sarcopterygii | ||
| The Vertebrates | Actinistia |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Teleostomi
├─Neopterygii
└─Sarcopterygii
├─┬─Onychodontiformes
│ └─Actinistia
│ ├─Miguashaia
│ └─┬─Diplocercides
│ └─┬─Hadronector
│ └─┬─Rhabdoderma
│ └─Coelacanthiformes
└─Rhipidistia
├─Dipnomorpha
└─┬─Rhizodontiformes
└─Osteolepiformes
├─Tristichopteridae
└─Elpistostegalia
├─Panderichthys
└─Tetrapoda
|
Contents
Overview |
Here we begin a series of pages on Actinistia, the coelacanth lineage. At the present time, we only cover Devonian and Carboniferous forms. If you care desperately about Mesozoic coelacanths, Peter Forey's 400+ page book is the Bible. This is the Spark Notes version of the Pentateuch.
At this point, it is traditional to do a little human interest story on Marjorie Courtenay-Latimer and the discovery of the living coelacanth, Latimeria. However, we have precious little interest in humans. Accordingly, we will direct the interested reader to the Wikipedia entry and links therein. It really is a tale worth hearing, but after one has read a dozen different versions of the story, it gets a bit old. As an aside, we should one day tell the story of Latimeria stories. It would be a study in itself to show how, over seventy years, judicious selections from the same pool of essential facts have been adapted as morality plays by interests as diverse as deconstructing feminists and unreconstructed Botha-style White Supremacists (and, we should add, also by the opponents of both). At any rate, except for a brief digression on the First Council of Nicea (325 A.D.), we will stick pretty much to fish.
Another subject we will not directly discuss is the origin of the Actinistia. At several points we have introduced our own take on the problem. In essence, 95% of all thinking on the origin of Actinistia was done before the discovery of truly basal forms like Psarolepis. The evolution of Actinistia needs to be completely re-evaluated in light of our new understanding of the primitive condition. We suspect that actinistians did not suddenly appear in the Middle Devonian with a bundle of completely unique characteristics. Rather many actinistian features (e.g. the structure of the cheek) may be primitive, or only slightly derived from the basal sarcopterygian condition.
Our treatment begins, in the usual haphazard fashion, with a series of random observations on Diplocercides and the relations between braincase and palate in the Sarcopterygii. From there, we pause only briefly at Euporosteus to consider a critical transition in cranial structure between the earliest known actinistian, Miguashaia, and the rest of the tribe. At this point, things fall apart completely as we take up the shattered remains of the possible actinistian Holopterygius. This gives us an opportunity to detour into the twisted topic of "relative warps analysis." Returning to fish, we next move to Hadronector and the contentious issues of sensory lines and the organization of the dermal bones of the actinistian skull table. Admittedly, we do not actually make an attempt to summarize the composition of the skull until we arrive at a long, meandering essay on Rhabdoderma. It is there that we finally have something to say about the plesiomorphy of the actinistians and the non-homology of most actinistian skull elements with those of the tetrapods. In addition, we include uncommented descriptions of a number of Bear Creek coelacanths: Caridosuctor, Polyosteorhynchus, and the incomparably odd Allenypterus.
Actinistia: (Coelacanthamorpha)
from the Middle Devonian. Group had considerable diversity during Carboniferous.
Sarcopterygii ::: Onychodontiformes + * : Miguashaia + (Diplocercides + (Euporosteus + (Hadronector + ((Allenypterus + Holopterygius) + (Lochmocercus + (Polyosteorhynchus + (Caridosuctor + (Rhabdoderma + Coelacanthiformes)))))))).
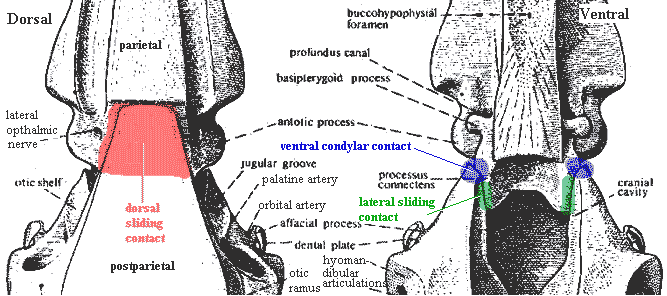
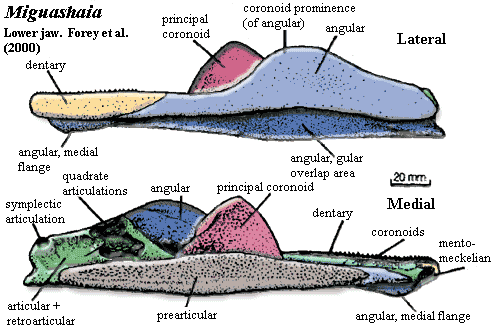
Characters: Rostrum: premaxilla small [J96]; maxilla absent [J96] [ZY02$] reversal); rostrum often broken up into numerous, non-standard dermal bones; often 1+ median unpaired rostrals [F98]; electroreceptive (probably) rostral organ morphologically similar to nasal capsules, located dorsomedially to nasal capsules [J80]; rostral 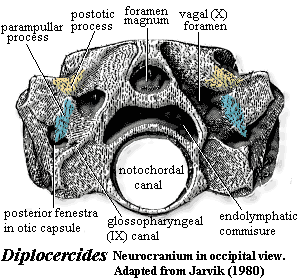 organ with paired anterior openings anterior to both nares and two pairs of posterior openings posterior to both nares [J80]; Skull Table: nomenclature (and comparability) of dermal skull table and surrounding area hopelessly confused -- see Rhabdoderma for discussion and tentative conclusions; two pairs of parietals, with anterior normally overlapping posterior or suturing irregularly [F98]; middle and posterior pit-lines near middle of postparietal [ZY02$]; infraorbital canal runs through postparietal [ZY02$]; spiracle and spiracular groove absent [J80]; Cheek: kinetic skull in cheek area; very large opercular [J96]; Braincase: braincase completely divided into otoccipital & sphenethmoid parts [F98] tendency for otoccipital to break up into separate ossification centers; notochord persistent, extending to rest against pit on posterior of sphenethmoid [F98]; otoccipital region short, with "afacial process" bearing toothplate & projecting laterally just ventral to otic shelf [J80] (see images at Diplocercides); otic region with posterior process from region of semicircular canals which bridges to region of exoccipitals (= parampular process) [J80]; endolymphatic duct opens on roof of otoccipital [J80]; endolymphatic commisure appears to join inner ear regions across occiput [J80]; vestibular fontanelle absent, but otic region has posterior foramen of unknown function [J80]; lateral commisure present on otoccipital with jugular canal and articulations for two-headed hyomandibular more or less as in osteolepiforms [J80] [F81]; afacial process located just ventral to small "prefacial eminence" on otic shelf [J80]; orbital artery probably passing between afacial process and prefacial eminence [J80]; intracranial joint with three elements [F81] [1]; basipterygoid process absent in more derived forms [J80]; deep lateral groove on dorsal sphenethmoid in orbitonasal area for supraorbital canal [LJ88]; nasal cavities separated from orbit by thick postnasal wall [J80]; solum nasi complete, no choana [J80]; nasal cavity with ventrolateral fossa for base of ligament connecting to autopalatine [J80]; Palate: parasphenoid ascending process absent [F98]; tripartite upper jaw (autopalatine, pterygoid, pterygoquadrate -- see pterygoid) [J80]; autopalatine with triangular dorsal ascending lamina for ligamentous attachment to ethmosphenoid, primitively in deep fossa ventrolateralis of nasal fossa [J80] [F98]; large triangular pterygoids,as contrasted with the more elongate and cleaver-like shape of other fishes [J96] [F+00]; often with loosely attached dermopalatines (usually 2) and ectopterygoid(s) [F98]; posterior margin of palatoquadrate erect or sloping backward [ZY02$]; pterygoquadrate with ascending process posterior to which
organ with paired anterior openings anterior to both nares and two pairs of posterior openings posterior to both nares [J80]; Skull Table: nomenclature (and comparability) of dermal skull table and surrounding area hopelessly confused -- see Rhabdoderma for discussion and tentative conclusions; two pairs of parietals, with anterior normally overlapping posterior or suturing irregularly [F98]; middle and posterior pit-lines near middle of postparietal [ZY02$]; infraorbital canal runs through postparietal [ZY02$]; spiracle and spiracular groove absent [J80]; Cheek: kinetic skull in cheek area; very large opercular [J96]; Braincase: braincase completely divided into otoccipital & sphenethmoid parts [F98] tendency for otoccipital to break up into separate ossification centers; notochord persistent, extending to rest against pit on posterior of sphenethmoid [F98]; otoccipital region short, with "afacial process" bearing toothplate & projecting laterally just ventral to otic shelf [J80] (see images at Diplocercides); otic region with posterior process from region of semicircular canals which bridges to region of exoccipitals (= parampular process) [J80]; endolymphatic duct opens on roof of otoccipital [J80]; endolymphatic commisure appears to join inner ear regions across occiput [J80]; vestibular fontanelle absent, but otic region has posterior foramen of unknown function [J80]; lateral commisure present on otoccipital with jugular canal and articulations for two-headed hyomandibular more or less as in osteolepiforms [J80] [F81]; afacial process located just ventral to small "prefacial eminence" on otic shelf [J80]; orbital artery probably passing between afacial process and prefacial eminence [J80]; intracranial joint with three elements [F81] [1]; basipterygoid process absent in more derived forms [J80]; deep lateral groove on dorsal sphenethmoid in orbitonasal area for supraorbital canal [LJ88]; nasal cavities separated from orbit by thick postnasal wall [J80]; solum nasi complete, no choana [J80]; nasal cavity with ventrolateral fossa for base of ligament connecting to autopalatine [J80]; Palate: parasphenoid ascending process absent [F98]; tripartite upper jaw (autopalatine, pterygoid, pterygoquadrate -- see pterygoid) [J80]; autopalatine with triangular dorsal ascending lamina for ligamentous attachment to ethmosphenoid, primitively in deep fossa ventrolateralis of nasal fossa [J80] [F98]; large triangular pterygoids,as contrasted with the more elongate and cleaver-like shape of other fishes [J96] [F+00]; often with loosely attached dermopalatines (usually 2) and ectopterygoid(s) [F98]; posterior margin of palatoquadrate erect or sloping backward [ZY02$]; pterygoquadrate with ascending process posterior to which 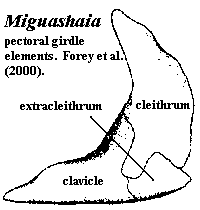 the antotic process (and the more anterior suprapterygoid process, if present) of the sphenethmoid articulates [J80] [F98]; supratemporal articulation absent [F98]; quadrate usually stout, with double condyle [F98]; Lower jaw see image at Miguashaia): dentary short [J96] [ZY02$]; dentary less than 1/2 length of jaw [F+00]; 4-5 coronoids [ZY02]; enlarge principle coronoid, triangular to quadrangular, often with anterior tilt [F98]; 2 large infradentaries; no infradentaries behind splenial [F+00]; surangular absent [ZY02$] reversal); 2 enlarged infradentaries (angular and splenial) [J96] [F98]; postsplenial: absent [ZY02$] (reversal); angular large, with raised crest [F98] [F+00] [ZY02$]; odd jaw hinge involving articulation with hyomandibular on a separate (in Mesozoic forms) retroarticular ossification of Meckel's cartilage through symplectic & stylohyal bones [J80] [F+00]; complex articular + retroarticular [F+00]; branchiostegal rays absent; Dentition: reduced marginal dentition; parasymphysial tooth whorl absent [ZY02$] reversal); dentary tooth row reaches anterior end of dentary [ZY02$] reversal); hyomandibula short and thick and does not articulate with palatoquadrate; Branchial hyomandibular very short and block-like, not attached to palate [F98]; hyomandibula articulates with a short interhyal or stylohyal [F98]; ceratohyal articulates ventrally with (a) ceratohyal ( leading to basibranchials) and (b) to autapomorphic symplectic (leading to jaw) [F98]; urohyal with broad posterior "wings" on either side of deep groove bordered by prominent ridges [LJ88]; branchial arches 1-3 typically with paired and/or separate basibranchials [F81]; Axial: no ossified centra, with arch elements having perichondral ossification [F98]; notochord broad, tapering rapidly in tail region [F98]; vertebrae short, with >100 in derived forms [F98]; almost all neural arches co-ossified with spines [F98]; supraneurals absent except for dorsal radials (epineurals) of caudal fin [F98]; fins other than caudal usually with fin lobes [J96]; dorsals supported by single basal; unlobed 1st dorsal common; 1st dorsal located anteriorly, often far anterior [F98]; 1st dorsal typically consisting of small number of unbranched rays supported directly on fin basal [F98]; basal of 1st dorsal with thickened ridges [F98] [2]; endoskeleton of 2nd dorsal and anal fins similar to paired fins, with 2+ radials, one radials producing mesomeres wich support fin rays [J96] [F98]; 2nd dorsal and anal fin mesomeres with dorsal & ventral processes similar to paired fin with 4 mesomeres [F98]; 2nd dorsal & anal fins almost chiral pair [F98]; caudal fin primitively epicercal with large webbed lower section, but in more derived forms diphycercal and 3-lobed, with middle section representing extension of notochord bearing a small tuft of lepidotrichia [J96] [F98]; pectoral fin with extracleithrum lying along posterior margin of cleithrum [F+00$]; Appendicular: each member of paired fins moves independently, so that motion converges with tetrapod walking; pectoral girdle not bound to skull, with supracleithrum and posttemporal absent [F98]; pectoral fin insertion at 50-70% body depth, with fin oriented vertically [F98]; pectoral girdle thin [F98]; anocleithrum unornamented, receives muscle from 4th branchial arch [F98]; extracleithrum present, usually attached to both clavicle & cleithrum [J96] [F98$]; pelvic fin insertion at about 50% body depth, behind 1st dorsal, with fins oriented horizontally [F98]; pelvic fin articulates with acetabulum on posterior basal [F98]; Other: large, rounded scales [J96]; heart elongate & tube-like [J96]; in some basal forms, abdomen contains long calcified (air?) sac or fat-filled lung (Latimeria); ovaviviparous.
the antotic process (and the more anterior suprapterygoid process, if present) of the sphenethmoid articulates [J80] [F98]; supratemporal articulation absent [F98]; quadrate usually stout, with double condyle [F98]; Lower jaw see image at Miguashaia): dentary short [J96] [ZY02$]; dentary less than 1/2 length of jaw [F+00]; 4-5 coronoids [ZY02]; enlarge principle coronoid, triangular to quadrangular, often with anterior tilt [F98]; 2 large infradentaries; no infradentaries behind splenial [F+00]; surangular absent [ZY02$] reversal); 2 enlarged infradentaries (angular and splenial) [J96] [F98]; postsplenial: absent [ZY02$] (reversal); angular large, with raised crest [F98] [F+00] [ZY02$]; odd jaw hinge involving articulation with hyomandibular on a separate (in Mesozoic forms) retroarticular ossification of Meckel's cartilage through symplectic & stylohyal bones [J80] [F+00]; complex articular + retroarticular [F+00]; branchiostegal rays absent; Dentition: reduced marginal dentition; parasymphysial tooth whorl absent [ZY02$] reversal); dentary tooth row reaches anterior end of dentary [ZY02$] reversal); hyomandibula short and thick and does not articulate with palatoquadrate; Branchial hyomandibular very short and block-like, not attached to palate [F98]; hyomandibula articulates with a short interhyal or stylohyal [F98]; ceratohyal articulates ventrally with (a) ceratohyal ( leading to basibranchials) and (b) to autapomorphic symplectic (leading to jaw) [F98]; urohyal with broad posterior "wings" on either side of deep groove bordered by prominent ridges [LJ88]; branchial arches 1-3 typically with paired and/or separate basibranchials [F81]; Axial: no ossified centra, with arch elements having perichondral ossification [F98]; notochord broad, tapering rapidly in tail region [F98]; vertebrae short, with >100 in derived forms [F98]; almost all neural arches co-ossified with spines [F98]; supraneurals absent except for dorsal radials (epineurals) of caudal fin [F98]; fins other than caudal usually with fin lobes [J96]; dorsals supported by single basal; unlobed 1st dorsal common; 1st dorsal located anteriorly, often far anterior [F98]; 1st dorsal typically consisting of small number of unbranched rays supported directly on fin basal [F98]; basal of 1st dorsal with thickened ridges [F98] [2]; endoskeleton of 2nd dorsal and anal fins similar to paired fins, with 2+ radials, one radials producing mesomeres wich support fin rays [J96] [F98]; 2nd dorsal and anal fin mesomeres with dorsal & ventral processes similar to paired fin with 4 mesomeres [F98]; 2nd dorsal & anal fins almost chiral pair [F98]; caudal fin primitively epicercal with large webbed lower section, but in more derived forms diphycercal and 3-lobed, with middle section representing extension of notochord bearing a small tuft of lepidotrichia [J96] [F98]; pectoral fin with extracleithrum lying along posterior margin of cleithrum [F+00$]; Appendicular: each member of paired fins moves independently, so that motion converges with tetrapod walking; pectoral girdle not bound to skull, with supracleithrum and posttemporal absent [F98]; pectoral fin insertion at 50-70% body depth, with fin oriented vertically [F98]; pectoral girdle thin [F98]; anocleithrum unornamented, receives muscle from 4th branchial arch [F98]; extracleithrum present, usually attached to both clavicle & cleithrum [J96] [F98$]; pelvic fin insertion at about 50% body depth, behind 1st dorsal, with fins oriented horizontally [F98]; pelvic fin articulates with acetabulum on posterior basal [F98]; Other: large, rounded scales [J96]; heart elongate & tube-like [J96]; in some basal forms, abdomen contains long calcified (air?) sac or fat-filled lung (Latimeria); ovaviviparous.
 Links: Coelacanth; Animal Diversity Web: Latimeria chalumnae (Coelacanth): Narrative; World Conservation Monitoring Centre - WCMC/WWF Species Under Threat; The Century in Review: Coelacanths; Basal Sarcopterygians; Coelacanth; pick.html; Actinistia a.k.a. coelacanths); Coelacanthinimorpha;
15sorh.pdf;
Animal Diversity Web: Actinistia; Molecular Phylogenetic Information on the Identity of the Closest ...; v1fish; Higher Ribosomal RNA Substitution Rates in Bacillariophyceae and; Devonian Times - More about Lobe-Fin Fishes; Close tetrapod relationships of the coelacanth Latimeria ...; Link to NCBI Front Page; Die evolutive Transformation der Oticalregion der Sarcopterygii ....
Links: Coelacanth; Animal Diversity Web: Latimeria chalumnae (Coelacanth): Narrative; World Conservation Monitoring Centre - WCMC/WWF Species Under Threat; The Century in Review: Coelacanths; Basal Sarcopterygians; Coelacanth; pick.html; Actinistia a.k.a. coelacanths); Coelacanthinimorpha;
15sorh.pdf;
Animal Diversity Web: Actinistia; Molecular Phylogenetic Information on the Identity of the Closest ...; v1fish; Higher Ribosomal RNA Substitution Rates in Bacillariophyceae and; Devonian Times - More about Lobe-Fin Fishes; Close tetrapod relationships of the coelacanth Latimeria ...; Link to NCBI Front Page; Die evolutive Transformation der Oticalregion der Sarcopterygii ....
Notes: [1] "The coelacanth intracranial joint has three basic skeletal components: dorsally there is a sliding portion between postero-ventral processes of the [parietals] and the undersurface of the [postparietals]; ventrally there is an articulation between the sphenoid condyles on the posterior face of the basisphenoid and the anterior face of the [prootic]; laterally there is a further sliding joint between the processus connectens of the basisphenoid and a groove on the inside of the prootic." [F81: 205]. [2] incorporated radials?
Carroll 1988); Forey 1981) [F81]; Forey 1998) [F98]; Forey et al. (2000) [F+00]; Janvier 1996) [J96]; Jarvik 1980) [J80]; Lelièvre & Janvier (1988) [LJ88]; Long 1995); Zhu & Yu (2002) [ZY02].
ATW060119.
Miguashaia: Schultze, 1973. M. bureaui Schultze, 1973 [4]; M. grossi Forey et al., 2000. May be closely related to Gavinia [F+00].
Range: Middle Devonian (Givetian) to Late Devonian Frasnian) of Baltics & North America.
 Phylogeny: Actinistia : (Diplocercides + (Euporosteus + (Hadronector + ((Allenypterus + Holopterygius) + Lochmocercus + (Polyosteorhynchus + (Caridosuctor + (Rhabdoderma + Coelacanthiformes))))))) + *.
Phylogeny: Actinistia : (Diplocercides + (Euporosteus + (Hadronector + ((Allenypterus + Holopterygius) + Lochmocercus + (Polyosteorhynchus + (Caridosuctor + (Rhabdoderma + Coelacanthiformes))))))) + *.
Characters: skull roof: skull roof long & narrow [F98]; snout may have had mosaic of small bones, but poorly known [F98]; two pairs of parietals [S73]; one parietal overlies entire orbit [F98]; tectal series with at least five elements, four over orbit [F98]; supraorbital canal runs through centers of tectals, rather than between tectals & skull table [F98]; skull table constricted at intracranial joint [F98]; postparietals long & narrow, expanding posteriorly [F98]; supratemporal present, with two intertemporals anterior to it (or intertemporal + dermosphenotic) [F98]; at least one pair of extrascapulars [F98]; cheek:cheek region extremely long [S73] [F98]; cheek bones closely sutured & may overlap slightly [F98]; orbit small & anterior, with eye oriented obliquely [F98]; lacrimojugal broadly contacts postorbital & squamosal [F98]; large gap between postorbital and supraorbital series of dermal bones [F+00] questioned in [F98]); postorbital large [F98]; squamosal large & elongate [S73] [F98]; opercular almost square [S73]; subopercular (dorsoventrally) narrow, located directly ventral to opercular [S73] [F98]; opercular & subopercular overlap shoulder girdle [S73]; gular very broad, largest bone in skull, suggesting head had broad ventral surface [S73] [F+00]; gular narrower anteriorly, with anterior recess [S73]; pterygoid elongate, with slightly concave ventral margin [S73] [3]; pterygoid ("entopterygoid") posterior margin flat & angled anterodorsally, with quadrate articulation in the posteroventral recess [S73]; pterygoid with fold or process mid-way along dorsal margin [F+00]; Lower jaw: lower jaw long and low [F+00]; dentary length 1/3 length of jaw [F+00]; no hook-shaped process on dentary [F+00]; dentary pore absent [F+00]; splenial small [F98]; large angular with coronoid prominence [F+00] [1]; angular with dorsal unornamented area and wide ventral overlap surface for gular [F+00]; principle coronoid triangular and inclined anteriorly [F98]; prearticular shallow [F+00]; articular fused with retroarticular [F+00]; retroarticular with strong process & distinct cartilage-capped articulation for symplectic [F+00]; retroarticular deep [F+00]; Dentition: premaxilla 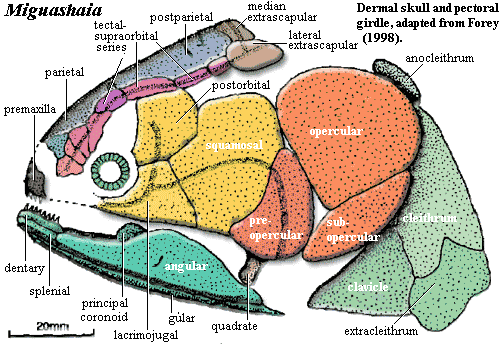 and dentary bearing strongly recurved teeth [S73]; ~11 teeth ankylosed to dentary [F98] [F+00]; epibranchials with long, slim denticles, not arranged in clusters [S73]; hypobranchials with denticles arranged in rows [S73]; Axial: neural & hemal fins & spines very broad distally and may abut one another [F98]; unpaired fins without fin lobes, except for upper caudal fin lobe [S73] (contra [F98]: anal fin may have small lobe); lepidotrichia in all fins segmented and distally bifurcated in unpaired fins (bifurcation in paired fins could not be determined) [S73]; 2nd dorsal & anal placed well posteriorly [F98]; caudal fin rays/radials >> 1:1 [F98]; caudal fin heterocercal, with upper lobe scaled, containing continuation of notochord and well-separated (not continuous with) conventional lower lobe [S73]; fin rays in upper lobe shorter [S73]; "caudal lepidotrichia in which successive segments interlock by means of step-like sutures" [F+00]; branched fin rays [F+00]; Appendicular: pectoral girdle broad [F+00]; all pectoral girdle bones ornamented with ridges [S73]; cleithrum with broad overlap areas on outer face for clavicle and extracleithrum, but no medial (meaning inner? anterior?) overlap regions [F+00]; extracleithrum nearly square [F+00]; extracleithrum both over- & underlaps cleithrum [F+00]; extracleithrum drawn out to point posteriorly [F+00] [2]; clavicle large and probably closely associated with antimere (primitive for Sarcopterygii, but unique in coelacanths) [F+00]; clavicle overlaps extracleithrum [F+00]; clavicle inturned anteriorly to form postbranchial lamina [F+00]; exposed interclavicle may be present, but isolated specimen may not be from same genus [F+00]; scapulocoracoid cartilaginous (since no trace of attachment on inner face of cleithrum) [S73] [F+00]; paired fins with narrow, relatively long fin lobes [S73]; scales with external ornament of tubercles running into ridges & fine chevron markings, and covered areas with radiating lines of spoon-shaped tubercles [F+00] (contra [F98]: ornament highly variable).
and dentary bearing strongly recurved teeth [S73]; ~11 teeth ankylosed to dentary [F98] [F+00]; epibranchials with long, slim denticles, not arranged in clusters [S73]; hypobranchials with denticles arranged in rows [S73]; Axial: neural & hemal fins & spines very broad distally and may abut one another [F98]; unpaired fins without fin lobes, except for upper caudal fin lobe [S73] (contra [F98]: anal fin may have small lobe); lepidotrichia in all fins segmented and distally bifurcated in unpaired fins (bifurcation in paired fins could not be determined) [S73]; 2nd dorsal & anal placed well posteriorly [F98]; caudal fin rays/radials >> 1:1 [F98]; caudal fin heterocercal, with upper lobe scaled, containing continuation of notochord and well-separated (not continuous with) conventional lower lobe [S73]; fin rays in upper lobe shorter [S73]; "caudal lepidotrichia in which successive segments interlock by means of step-like sutures" [F+00]; branched fin rays [F+00]; Appendicular: pectoral girdle broad [F+00]; all pectoral girdle bones ornamented with ridges [S73]; cleithrum with broad overlap areas on outer face for clavicle and extracleithrum, but no medial (meaning inner? anterior?) overlap regions [F+00]; extracleithrum nearly square [F+00]; extracleithrum both over- & underlaps cleithrum [F+00]; extracleithrum drawn out to point posteriorly [F+00] [2]; clavicle large and probably closely associated with antimere (primitive for Sarcopterygii, but unique in coelacanths) [F+00]; clavicle overlaps extracleithrum [F+00]; clavicle inturned anteriorly to form postbranchial lamina [F+00]; exposed interclavicle may be present, but isolated specimen may not be from same genus [F+00]; scapulocoracoid cartilaginous (since no trace of attachment on inner face of cleithrum) [S73] [F+00]; paired fins with narrow, relatively long fin lobes [S73]; scales with external ornament of tubercles running into ridges & fine chevron markings, and covered areas with radiating lines of spoon-shaped tubercles [F+00] (contra [F98]: ornament highly variable).
Notes: [1] [F+00] list a large number of additional jaw characters which, as they note, are also plesiomorphic. See Actinistia. [2] This might suggest that the extracleithrum is a remnant of the pectoral fin spine, and thus primitive for Sarcopterygii, or even Gnathostomata. [3] our very limited German is not really up to this job, and the translation is uncertain. The original is mit seinem leicht dorsal geschwungenen Ventralrand. [4] Schultze [S73] notes that the only known specimen of M. bureaui is a juvenile or subadult.
References: Forey 1998) [F98]; Forey et al. (2000) [F+00]; Schultze (1973) [S73]. ATW060114.
checked ATW041103