Mammalia: Triconodonta
Taxa on This Page
- Amphilestes X
- Gobiconodon X
- Jeholodens X
- Triconodonta X
Triconodonts: Cusps & Confusion
Like so many things, it all seemed simple at first. Therapsids were "reptiles" with reptile slicing and tearing teeth. Cynodonts were reptiles, but had some rather randomly placed enameloid pustules on the crowns of the teeth, sometimes with a circular arrangement of smaller cusps along the outside. This allowed some crushing and perhaps a bit of shearing. The triconodonts came next. They got things properly organized by putting three main cusps in a straight line, with or without outer cingula and accessory cusps. This radically improved shearing. The symmetrodonts moved the central cusp outward or inward to create a triangle pattern, a proto-trigon, with even better shearing and some grinding in the interstices and corners. Finally (blast of trumpets) the true tribosphenic molar arrived, which allowed grinding on a trigonid basin, as well as all of the previously noted cutting, shearing, ripping, slicing, tearing, gnashing, gouging, and so on.
Like so many things, it was too linear to last. Symmetrodonts, triconodonts, and even tribosphenics are now in utter confusion. There would be little point to tracing all the steps in this gradual descent into chaos. Generally speaking, the seemingly linear array of mammaliforms was an artifact of preservation. Until recently, we had little but teeth to look at between the Triassic and the Paleocene. That gap is slowly filling with bits and pieces of other things in a very irregular way. The linear arrangement was also an artifact of geography. Before the 1990's almost all the mammaliform material came from Europe and North America. New and better material from China, and also Australia, Africa and India, failed to fit the pattern.
It seemed the triconodonts were headed towards the same doom. In the 1920's Simpson wrote extensively on the Triconodonta, a group which he conceived to include many genera we would, today, refer to as symmetrodonts. Over the next few decades, Triconodonta shed, first its symmetrodont members, then the Triassic Morganucodon and others we might refer to as docodonts. With the application of cladistic techniques, the monophyly of the remaining triconodonts became suspect. Most of the triconodont genera outside the Triconodontidae show up as a series of unrelated branches off the main stem of mammaliform evolution -- either just outside or just inside the crown group mammals. Later, even the monophyly of the Triconodontidae was (and is) questioned. There is increasing evidence that the Jurassic Priacodon, Trioracodon, and Triconodon are rather different from the Cretaceous North American forms. Cifelli & Madsden (1998) have proposed that the difference are large enough to suspect that the two generations of Triconodontidae are polyphyletic. That is, their last common ancestor lies outside the Triconodontidae as currently understood.
However, the most recent analysis gives some reason to believe that there is a solid core of truth to the Triconodonta. Luo et al. (2002) find surprisingly robust support for at least some kind of Triconodonta -- some clade of triconodonts more inclusive than just the Triconodontidae. Since the deconstruction of this taxon may have been premature, it will be treated as a clade for the present. APW
040806.
So What's a Triconodont?
 I'm so glad you asked. A lot of the history and description of triconodonts is still dental. For that reason, it may be useful to refer to the painfully crude figure at right. As we will have to get into this stuff in absurd detail at some other points, we eschew the fine points of chewing for the moment.
I'm so glad you asked. A lot of the history and description of triconodonts is still dental. For that reason, it may be useful to refer to the painfully crude figure at right. As we will have to get into this stuff in absurd detail at some other points, we eschew the fine points of chewing for the moment.
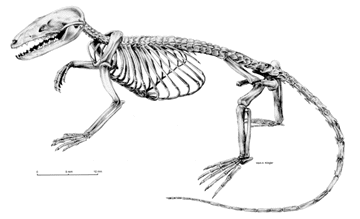
In overall aspect and lifestyle the triconodonts resembled a sort of small Mesozoic raccoon or possum. The "small" is by Cenozoic standards. By comparison to other Mesozoic mammaliforms, the triconodonts were giants. Some probably exceeded 50 cm in length and weighed in at over 1 kg. Their dentition was rather unspecialized and versatile, suggesting an omnivorous lifestyle. Jugulator may have been primarily carnivorous. Cifelli & Madsden (1998). Two poorly-known Moroccan genera, Dyskritodon and Ichthyoconodon were piscivorous, perhaps even aquatic. Sigogneau-Russell 1995). The very odd hind limbs of Gobiconodon Jenkins & Schaff, 1988) suggest that it also had some unusual lifestyle, although we cannot tell what that may have been. The sheer diversity of triconodont remains in some North American locations requires that we assume some degree of specialization. However, the general impression left by most triconodonts is that they were generalist omnivorous foragers.
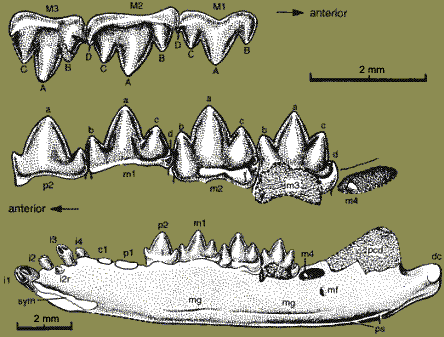
The first lower incisor was procumbent and large, sometimes very large, with deep roots extending far back under the other premolar teeth. At some point in early triconodont evolution, the canines were converted to premolariform teeth. However, the second incisors are generally caniniform, sometimes strikingly so. At least in the case of Gobiconodon, the animal may also have had deciduous ("baby") premolar teeth which were not replaced in adulthood. These peculiarities have caused a good deal of confusion and contradiction in the tooth literature. Not only are the tooth morphologies misleading, but there are odd gaps in the tooth row which do not correspond to the usual post-canine diastemae. The published dental formulae are often inconsistent and misleading -- and sometimes just plain wrong.
The premolars are strongly molariform. The molars tend to interlock with their mesial and distal neighbors in ways which become more elaborate and precise in later forms. The advantage of this unusual adaptation may relate to the triconodont dental strategy. The unworn molar cusps are smooth and conical. Sharp points and cutting edges were produced by wear, rather than genetics. Generally, the lower molar cusps occluded slightly medial to the upper cusps and sheared down the medial face. Consequently, the molars were constantly subject to considerable lateral stresses. The interlocking mechanism stabilized the molars, putting the lateral forces to work to scrape sharp, regular edges on the molar cusps rather than simply loosening the teeth over time.
The dentition was backed up with powerful masseter muscles which occupied a large, deep masseteric fossa on the lateral side of the dentary. Most triconodonts have a foramen leading inward from the anteroventral corner of the masseteric fossa. The tooth row is frequently shortened: pushed forward, as it were, by the growth of the enormous coronoids bearing the masseteric fossae. What is known of the rest of the skull is -- not much. There are a two triconodonts known from substantially complete skulls: Jeholodens and Gobiconodon, but there is little about either which seems unusual or even informative. The jugal is perhaps unusually constrained, the maxilla correspondingly flamboyant. This suggests a relationship with the multituberculates, which may or may not be the case. The triconodont lacrimal is also large, but it is small or absent altogether in many multituberculates. The braincase is known in part, and the petrosal of Trioracodon is discussed in a separate essay. In essence it can be conceived as basal mammaliform braincase stretched to its absolute limits by an expanded brain.
The post-cranial skeleton is known only from Jeholodens, Gobiconodon, and (in small part) from Priacodon. It is probably unsafe to generalize from such a small sample, but we will do so for lack of better information. As one might guess, the post-cranial skeleton is a mosaic of basal mammaliform and therian characters. The atlas-axis complex is quite mammalian, but cervical ribs are sometimes found. The thoracic vertebrae bear rather long neural spines, sharply slanted backwards. Intervertebral articulations are found both on the centrum (pre- and postzygapophyses) and on the arches (metapophyses and anapophyses). The thoracic ribs are long and relatively uncurved distally, suggesting a deep body. The lumbar region is notably distinct from the thoracic domain, but the transition is more gradual than in modern mammals. The lumbar vertebrae look normal, but have small ribs which are not fused to the centra. The tail is poorly known, but probably rather thin and short.
The triconodont pectoral girdle is quite modern-looking. In particular, the scapular spine is prominent, and the supra- & infra-spinous fossae are well-developed. The acromion is hook-like and extends below the glenoid as in modern mammals. The humeral articulation is complex, but appears to be generally similar to modern mammals and faces posteroventrally. However, the humerus itself is an antique. The shaft is twisted about 30°, and has an extensive deltopectoral crest, as well as a probable latissimus dorsi crest. The distal humerus bears well-developed epicondyles, and even an entepicondylar foramen. The ulnar articulation is quite strange. Anteriorly, it is a condyle -- the basal character state -- but posteriorly, it takes the form of a trochlea as in therian mammals. This odd character state is of obvious interest in finding the phylogenetic place of triconodonts. Unfortunately, the ulnar articulation is known only for a few taxa. morganucodontids, Cimolodonta derived multituberculates), and the platypus Ornithorhynchus have the primitive condyle. The hybrid state is found in triconodonts, the symmetrodont Zhangheotherium and a few transitional forms (Vincelestes, Henkelotherium). Even more interesting, a similar transitional state is found for the radial articulation only in Zhangheotherium and the two transitional mammals. Triconodonts, monotremes and multituberculates have the primitive condylar arrangement for the radius. Ji et al. (2002). The pelvic girdle is much less well-known and certainly less informative. In general it is therian. The femur of Gobiconodon is peculiar in detail, if pedestrian (literally!) in overall design. It comes with unidentified trochanters and a weirdly-formed lesser trochanter that suggest some unique postural adaptation. APW 040806.
Triconodont Phylogeny: On Luo Blows and Cingular Distinctions
The description above seems generally consistent with the loose consensus developing around the results of Luo et al. 2002). In brief, this outline of mammaliform phylogeny is that the first reasonably clear mammaliform grouping are the docodonts, including Morganucodon. These are followed by Hadrocodium and some early symmetrodont-like things, notably Kuehneotherium. The crown group Mammalia are next, with the monotremes splitting off. After that, there lies a chaotic mixture of more symmetrodonts and triconodonts, which probably includes bona fide clades of "acute angle symmetrodonts" and an augmented Triconodontidae, i.e. something more inclusive than Triconodontidae, but less inclusive than the traditional Triconodonta. Finally, we arrive at the non-tribosphenic proto-therians (like Vincelestes and Peramus), followed by the Theria, and close cousins with tribosphenic molars. In extremely simplified form:
Cynodonts
├─Docodonta & Morganucodon (clade)
└─┬─Hadrocodium, Kuehneotherium (plesion)
└─Mammalia
├─Monotremata (clade)
└─┬─Triconodonta (plesion, but including a large clade)
├─"acute angle symmetrodonts" (plesion including some kind of clade)
└─┬─non-tribosphenic proto-therians
└─Tribosphenida (= Boreosphenida)
There are, however, two kinks in this vague but smooth succession. One is the Allotheria (haramyids and multituberculates). Luo and coworkers find that Haramiyavia is not even a mammaliform but an aberrant cynodont. The multituberculates are found to be true mammals, emerging from the same chaotic region that produces the triconodonts and symmetrodonts. Nevertheless, Luo et al. confess that their analysis cannot confidently eliminate the possibility of a monophyletic Allotheria. Given how far Haramiyavia lies from the multituberculates in their original cladogram, one may reasonably become a bit suspicious.
The second problem is the Australosphenida. Rich and others have shown that mammaliforms with real tribosphenic molars, the Ausktribosphenidae, were present in Australia as early as the middle Cretaceous. Rich et al. (2001). This is sensible, as the monotremes had to come from somewhere. However, if the tribosphenic molar were present this early in the game, where did this mass of vehemently non-tribosphenic symmetrodonts and triconodonts come from between the monotremes and the therians? To solve this conundrum, Luo, Cifelli and Kielan-Jaworowska proposed that the tribosphenic molar evolved twice, independently. Luo et al. (2001a).
In the main, these issues are taken up elsewhere. However, they depend on a close reading of the dental characters; and, for that reason, it seemed appropriate to look closely at the treatment of dental characters in Luo et al. 2002). While we are strongly inclined to agree with the results of these authors, their treatment of dental characters is not encouraging. We won't beat this issue to death. It would require an excruciatingly long and tedious essay to point out the many problems, and -- frankly -- the value of our opinions on the subject doesn't justify the effort required to express them. A single example will do.
A substantial number of dental characters are included which are not applicable to animals without certain minimal dental equipment: trigon, protocone, or what have you. The only correct way to score these, or any other, characters is to begin with the outgroup condition as plesiomorphic. If that means "not applicable" is scored as zero, that's perfectly OK. What is not permissible is to score "not applicable" as if it were missing data. This is what Luo et al. do. The parsimony algorithms used in PAUP don't treat missing data as missing. They fill in the blank with whatever state will minimize the number of character transformations required. This has important consequences. Note that the triconodonts are bracketed between Australosphenida and stem boreosphenids, both of which have tribosphenic molars. If triconodonts are scored '0' for not applicable, two extra character state changes are required to get from australosphenids to triconodonts to boreosphenids. If triconodonts are scored '?' for not applicable, those steps are eliminated. PAUP will assign trigonids to triconodonts and no one will be the wiser. Bottom line: if the triconodonts had been properly scored, would they have been pushed out of this region and perhaps out of crown group Mammalia? Possibly. I would wager that the multituberculates definitely would be squeezed out of Mammalia. A very large fraction of Luo's dental characters are "not applicable" to multis and were scored incorrectly as a result. APW
040806.
Descriptions
 Triconodonta: = Eutriconodonta sensu [L+02], i.e. Amphilestes + Triconodon).
Jugulator, Triconodon, Repenomamus
Triconodonta: = Eutriconodonta sensu [L+02], i.e. Amphilestes + Triconodon).
Jugulator, Triconodon, Repenomamus
Range: Middle Jurassic (Late Triassic?) to Late Cretaceous of Laurasia & North Africa.
Carnivorous(?), up to cat-sized; septomaxilla present, without ventromedial shelf [L+02, 1]; lacrimal large, triangular & pointed anteriorly [L+02, 0]; tabular absent [L+02, 1]; anterior jugal excluded from anterior orbit & doesn't contact maxilla [L+02, 1]; squamosal doesn't participate in braincase [L+02, 0]; temporomandibular joint anterior to fenestra vestibuli (possible mammal synapomorphy) [L+02] [3]; glenoid on platform of arch & facing ventrally [L+02, 1]; zygomatic arch constricted between glenoid & cranial squamosal [L+02, 1]; post glenoid process present [L+02, 1]; distinct external auditory meatus [W+01]; no basisphenoid "wing" covering cochlear housing [L+02, 1]; post-dentary bones fully incorporated in middle ear [W+01]; petrosal forms large portion of ventral braincase, as well as lateral braincase through an anterior lamina; internal auditory meatus present as grove, but no cribiform foramina; [L+02, 2]; cochlea uncoiled, with long cylindrical promontorium [L+02, 1]; possibly shows major expansion of brain; bony secondary palate extending posterior to tooth row [L+02, 2]; post-dentary trough absent [L+02]; mandibular symphysis unfused [L+02]; dentary angular process unknown or absent; ventral surface of dentary rather straight; separate ossified Meckelian in some (may have functioned for attachment of m. pterygoidius medius) [W+01]; Meckelian groove weakly developed [L+02]; Meckelian groove convergent with ventral border of mandible (except Amphilestes) [L+02]; pterygoid fossa on medial face of jaw [L+02]; masseteric fossa ventral margin well-defined, thin ridge [L+02]; masseteric fossa with distinct crest along anterior border of coronoid process [L+02]; pterygoid shelf along ventral part of coronoid process, reaching condyle via low ridge {L+02]; dentary condyle is continuous with the semicircular posterior margin of dentary & faces dorsally due to upturning of posteriormost portion of dentary [L+02]; dentary peduncle ventral margin continuous with ventral margin of dentary [L+02]; dentary condyle at or below ventral level of post-canine alveoli [L+02]; coronoid absent (except Gobiconodon) [L+02]; splenial absent [L+02]; usually 3+ lower incisors (Gobiconodon with 2, Trioracodon unknown) [L+02]; large canines; main cusps of lower premolars in a straight line [L+02]; last lower premolar with distal cingulid cusp d [L+02]; lower premolars with lingual cingulid small or absent [L+02]; teeth occlude mostly with one tooth on opposite jaw; molars not replaced except in Gobiconodon [L+02]; lower molars with three main cusps in single row [L+02]; most with 4-5 lower molars {L+02]; lower molar cusps all somewhat laterally compressed [L+02]; cusps b & c of roughly equal size [L+02]; a-b cristid (paracristid) longitudinally oriented [L+02]; f cusp absent [S95]; mesial & distal transverse cingulids absent [L+02]; m3 is largest [L+02] [2]; last lower molar erupts medial to coronoid process [L+02]; upper molars with distinct, straight labial cingulum (no ectoflexus or cingular cusps) [L+02]; no grinding protocone or analog [L+02]; cuspule E/metastyle present (reversed in Triconodontidae) [L+02]; upper molars do not interlock [L+02]; long tail; independently evolved (?) therian-type shoulder girdle; primitive pelvis, with sprawling hind limbs; calcaneum with peroneal shelf & without peroneal process [L+02].
Note: [2] [L+02] score this character (their #59) in an odd fashion as discussed in the essay above. [3] In the text of the article, [L+02] indicate that the jaw joint is anterior to the fenestra vestibuli. However, in the character list, both Jeholodens and Gobiconodon are scored as having the joint lateral or posterior to the fenestra, i.e., char. 186 = 0.
Image: Jeholodens jenkinsi (Early Cretaceous of China) from the 1999 press release issued by the Carnegie Museum.
Links:
Untitled Document (questioning monophyly); ??[Khronologeeya Protsessov] (Russian); eslabon (Portuguese); La galerie de la page jurassique : la faune terrestre (French); Paleontology and Geology Glossary: T; Abstract of SVP 1997 JVP 17(3):54A; Mesozoic Mammals; Docodonta and Triconodonta, an internet directory; Carnegie Museum of Natural History News; Nature Science Update; pujaw.gif; Triconodonta.
References: Ji et al. (2002) [J+02]; Kielan-Jaworowska et al. (2000); Luo et al. (2001); Luo et al. 2002) [L+02]; Sigogneau-Russell 1995) [S95]; Wang et al. (2001) [W+01]. APW 040806.
 Gobiconodon: = Guchinodon) G. borissiaki Trofimov (1978); G. hoburensis Trofimov (1978); G. ostromi Jenkins & Schaff (1988). Closely related to Hangjinia chowi Godefroit (1999) and possibly Klamelia Chow & Rich (1984) (contra, [K+00]).
Gobiconodon: = Guchinodon) G. borissiaki Trofimov (1978); G. hoburensis Trofimov (1978); G. ostromi Jenkins & Schaff (1988). Closely related to Hangjinia chowi Godefroit (1999) and possibly Klamelia Chow & Rich (1984) (contra, [K+00]).
Range: Early Cretaceous II (Aptian or Albian) of Mongolia & North America.
Phylogeny: Triconodonta: (Amphilestes + (Jeholodens + Triconodontidae)) + *.
Characters: skull 3-10 cm [K+00]; septomaxilla present [JS88]; maxilla with 5 rounded fossae on palatal process [K+00]; frontal does not contact maxilla [J+02]; prominent sagittal crest [J+02]; jugal does not reach maxilla & does not participate in anterior of orbit [J+02]; cranial portion of squamosal narrow, does not participate in braincase [J+02]; squamosal with postglenoid depression on skull base [J+02]; foramen for superior ramus of stapedial artery enclosed [H+97]; glenoid faces ventrally [J+02]; squamosal with distinct postglenoid process [J+02]; dentary squamosal jaw joint only; cochlear housing promontorium) elongate & cylindrical [J+02]; cochlea short & uncoiled [J+02]; braincase without bulge at level of parietal region [J+02], [L+02, 0]; extensive symphysial region accommodating deep roots of incisors [JS88]; symphysial surfaces with numerous vascular foramina & so probable ligamentous, flexible symphysis [JS88]; distinctive mandibular foramen fully formed; Meckelian groove is vestigial or absent in adult [H+97] (contra [L+02] "weakly developed" but not vestigial or absent); bony secondary palate extending posterior to tooth row [J+02]; dentary angular process absent [J+99]; masseteric fossa deep [JS88]; dentary medial face, pterygoid
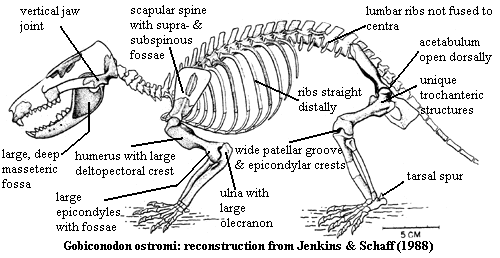 fossa present [J+02]; medial pterygoid ridge along ventral border of coronoid process & reaching condyle via low crest [J+02]; coronoid process inclined posteriorly [JS88] at 145° [J+02]; coronoid bone present [L+02] [JS88]; dentary condyle massive & bulbous, broad in dorsal view [J+02] and pisiform transversely? [JS88]; dentary condyle dorsally directed [JS88]; dentition dental formula 2/1, 1/1, 4+/3-4, 4-5/5 [JS88] [3]; I1 small [JS88]; small "crypt" between I1 & I2 [JS88]; I2 very large & caniniform, recurved with crest along distal surface [JS88]; 2 lower incisors [J+02]; palatal fossa medial to I2 for reception of i2 [JS88]; diastema follows I2 [JS88]; i1 long and procumbent [J+02] [JS88]; i1 root extending to level of p2! [JS88]; small upper & lower canines present, with single roots [J+02]; canines similar to premolars [JS88]; premolars 4/4 [J+02]; first upper premolar erect, with diastema [J+02] (unlikely -- see note [3]); p1-p3 with decreasing procumbency [K+00]; p2 & p3 trenchant and transversely compressed [JS88]; main cusp a of last p symmetrical [H+97]; premolar cusp a with sharp pre- & post-cristids, continuous with cristids on b & c cusps [JS88]; last lower premolar with b much smaller than c [L+02] [JS88]; last lower premolar with c distinct, but much shorter than a based on valley between them [L+02]; labial cingulid present in ultimate lower premolar [L+02]; last lower premolar with symmetrical central a cusp & labial cingulid [J+02]; molariform replacement, sequentially from mesial to distal [JS88]; "interlocking mechanism between lower molariforms in Gobiconodon is of the pattern seen in Kuehneotherium and Tinodon"; molar occlusion with a occluding in front of B and into the embrasure between the opposite upper tooth and the preceding upper tooth [J+02] (contra a occludes between C and D on posterior cingulum [K+02]); cusp A occludes with cusp b with strong shearing facet on mesiobuccal face of b [JS88]; A also occludes between cusps e & f [JS88]; B occludes with buccal surface of preceding c, & C with a [JS88]; occlusal pattern consistent with medial translation & rotation of mandible [JS88]; upper molar cusps have "incipient" triangular arrangement with increasing triangulation along molar row, A cusp being more lingual [K+00]; last lower molar cusps in a single row [J+02]; upper molars with increasing triangulation along molar row [J+02]; upper molars with B/paracone taller & wider than C/metacone [J+02]; M1 & m1 laterally compressed [J+02]; lower molar crowns rectangular in occlusal view [JS88]; cusps in a straight line, not triangulated [J+02] [JS88]; primary cusps tall & lanceolate [JS88]; cusp a dominates crown [S95]; cusp a with sharp pre- & post-cristids, continuous with cristids on b & c cusps [JS88]; lower molar cusps a>b=c>d [S95] [JS88]; c > b on m1-m2 & more robust on all lower molars [JS88]; b & c same height [J+02]; height of c <40% height of a [L+02]; lower molars with distinct lingual cingulid [J+02] (contra lingual cingulid incomplete on m1 & m5 [JS88]); lingual cingulum on most lower molars continues around mesial margin to mesiobuccal corner, with some individual variation [JS88] [4]!!; lower molars with mesiolingual & mesiobuccal cingular cuspules (e & f) [J+02]; talonid absent [J+02]; lower molars interlock, with d fitting between e & f cingulid cuspules of next more distal molar [J+02] [K+00]; axial skeleton atlas ribs fused in adults [H+97] (contra [J+02], who state this is not preserved); 13 thoracic vertebrae [J+02] (11+, probably 13 [JS88]); anterior thoracic vertebrae with elongate spines angled posteriorly at 50-55° [JS88]; pedicles of arch well anterior and deeply notched posteriorly [JS88]; zygapophyses indistinct, with articulation through arches [JS88]; some degree of intervertebral rotation permitted [JS88]; thoracic ribs with long, uncurved distal section, indicating deep body [JS88]; lumbar vertebrae with anteroposteriorly long blades, tilted back ~25° and elongate anapophyses [JS88]; lumbar ribs flat, spatulate not fused to centra [JS88]; pectoral
scapula medial surface flat [J+02]; scapula with acromion process hook-like and extending below glenoid [J+02] (as in therians, but not monotremes); acromion robust [JS88]; scapula with both supra & infraspinous fossae [JS88]; scapula with supraspinous fossa fully developed along entire dorsal border of scapula [J+02]; coracoid small & without posterior process [J+02]; (contra coracoid probably large [JS88]); procoracoid fused to sternum & absent as separate element [L+02]; glenoid articular surface at angle to scapula & faces mostly posterior (not laterally) [L+02]; glenoid rounded (not elongate) & concave not saddle-shaped) [L+02]; glenoid of scapula concave (instead of saddle shaped) and facing posteroventrally [H+97] [J+02]?; humeral head weakly inflected [J+02]; humeral shaft with strong torsion (>30°) [J+02]; humerus deltopectoral crest extending past midpoint of shaft [J+02] [JS88]; humerus with 2nd, medial crest (teres major or latissimus dorsi process) [JS88]; humerus epicondyles robust [L+02]; very broad epicondyles with large entepicondylar foramen [JS88]; humerus ulnar trochlea present posteriorly, with vestigial condyle anteriorly [J+02] [2]; capitulum hemispherical [JS88]; ulna proximal articulation oblique to shaft & appears to be condylar [JS88]; ulna olecranon robust & expanded proximally [JS88]; ulna sigmoid & mediolaterally narrow (typical mammal) [JS88]; ulna shaft anteroposteriorly broad [JS88]; both epipodials with flat distal articulations & no styloid process [JS88]; pelvic acetabular dorsal emargination closed [H+97] (contra, [J+02]) (contra [JS88], dorsal margin "broadly open 'V'"); pelvic elements unfused within acetabulum [J+02]; no femoral neck, femoral head oriented dorsally [J+02] (contra [JS88] femoral head set off by flat, broad neck); lesser trochanter present as broad flange with unique deep fossa dorsally [JS88]; additional flange-like trochanter distal to greater trochanter [JS88]; broad, shallow patellar groove flanked by crest-like epicondyles [JS88]; fibula does not contact distal femur [J+02] (compare [JS88] proximal fibula "probably" contacted tibia, not femur); external foot "spur" as in Zhangheotherium and monotremes [H+97] [L+02, 1] [JS88]; sesamoid bone present in the pedal flexor tendon [H+97] (contra [J+02]); metapodials & phalanges short & robust [JS88].
fossa present [J+02]; medial pterygoid ridge along ventral border of coronoid process & reaching condyle via low crest [J+02]; coronoid process inclined posteriorly [JS88] at 145° [J+02]; coronoid bone present [L+02] [JS88]; dentary condyle massive & bulbous, broad in dorsal view [J+02] and pisiform transversely? [JS88]; dentary condyle dorsally directed [JS88]; dentition dental formula 2/1, 1/1, 4+/3-4, 4-5/5 [JS88] [3]; I1 small [JS88]; small "crypt" between I1 & I2 [JS88]; I2 very large & caniniform, recurved with crest along distal surface [JS88]; 2 lower incisors [J+02]; palatal fossa medial to I2 for reception of i2 [JS88]; diastema follows I2 [JS88]; i1 long and procumbent [J+02] [JS88]; i1 root extending to level of p2! [JS88]; small upper & lower canines present, with single roots [J+02]; canines similar to premolars [JS88]; premolars 4/4 [J+02]; first upper premolar erect, with diastema [J+02] (unlikely -- see note [3]); p1-p3 with decreasing procumbency [K+00]; p2 & p3 trenchant and transversely compressed [JS88]; main cusp a of last p symmetrical [H+97]; premolar cusp a with sharp pre- & post-cristids, continuous with cristids on b & c cusps [JS88]; last lower premolar with b much smaller than c [L+02] [JS88]; last lower premolar with c distinct, but much shorter than a based on valley between them [L+02]; labial cingulid present in ultimate lower premolar [L+02]; last lower premolar with symmetrical central a cusp & labial cingulid [J+02]; molariform replacement, sequentially from mesial to distal [JS88]; "interlocking mechanism between lower molariforms in Gobiconodon is of the pattern seen in Kuehneotherium and Tinodon"; molar occlusion with a occluding in front of B and into the embrasure between the opposite upper tooth and the preceding upper tooth [J+02] (contra a occludes between C and D on posterior cingulum [K+02]); cusp A occludes with cusp b with strong shearing facet on mesiobuccal face of b [JS88]; A also occludes between cusps e & f [JS88]; B occludes with buccal surface of preceding c, & C with a [JS88]; occlusal pattern consistent with medial translation & rotation of mandible [JS88]; upper molar cusps have "incipient" triangular arrangement with increasing triangulation along molar row, A cusp being more lingual [K+00]; last lower molar cusps in a single row [J+02]; upper molars with increasing triangulation along molar row [J+02]; upper molars with B/paracone taller & wider than C/metacone [J+02]; M1 & m1 laterally compressed [J+02]; lower molar crowns rectangular in occlusal view [JS88]; cusps in a straight line, not triangulated [J+02] [JS88]; primary cusps tall & lanceolate [JS88]; cusp a dominates crown [S95]; cusp a with sharp pre- & post-cristids, continuous with cristids on b & c cusps [JS88]; lower molar cusps a>b=c>d [S95] [JS88]; c > b on m1-m2 & more robust on all lower molars [JS88]; b & c same height [J+02]; height of c <40% height of a [L+02]; lower molars with distinct lingual cingulid [J+02] (contra lingual cingulid incomplete on m1 & m5 [JS88]); lingual cingulum on most lower molars continues around mesial margin to mesiobuccal corner, with some individual variation [JS88] [4]!!; lower molars with mesiolingual & mesiobuccal cingular cuspules (e & f) [J+02]; talonid absent [J+02]; lower molars interlock, with d fitting between e & f cingulid cuspules of next more distal molar [J+02] [K+00]; axial skeleton atlas ribs fused in adults [H+97] (contra [J+02], who state this is not preserved); 13 thoracic vertebrae [J+02] (11+, probably 13 [JS88]); anterior thoracic vertebrae with elongate spines angled posteriorly at 50-55° [JS88]; pedicles of arch well anterior and deeply notched posteriorly [JS88]; zygapophyses indistinct, with articulation through arches [JS88]; some degree of intervertebral rotation permitted [JS88]; thoracic ribs with long, uncurved distal section, indicating deep body [JS88]; lumbar vertebrae with anteroposteriorly long blades, tilted back ~25° and elongate anapophyses [JS88]; lumbar ribs flat, spatulate not fused to centra [JS88]; pectoral
scapula medial surface flat [J+02]; scapula with acromion process hook-like and extending below glenoid [J+02] (as in therians, but not monotremes); acromion robust [JS88]; scapula with both supra & infraspinous fossae [JS88]; scapula with supraspinous fossa fully developed along entire dorsal border of scapula [J+02]; coracoid small & without posterior process [J+02]; (contra coracoid probably large [JS88]); procoracoid fused to sternum & absent as separate element [L+02]; glenoid articular surface at angle to scapula & faces mostly posterior (not laterally) [L+02]; glenoid rounded (not elongate) & concave not saddle-shaped) [L+02]; glenoid of scapula concave (instead of saddle shaped) and facing posteroventrally [H+97] [J+02]?; humeral head weakly inflected [J+02]; humeral shaft with strong torsion (>30°) [J+02]; humerus deltopectoral crest extending past midpoint of shaft [J+02] [JS88]; humerus with 2nd, medial crest (teres major or latissimus dorsi process) [JS88]; humerus epicondyles robust [L+02]; very broad epicondyles with large entepicondylar foramen [JS88]; humerus ulnar trochlea present posteriorly, with vestigial condyle anteriorly [J+02] [2]; capitulum hemispherical [JS88]; ulna proximal articulation oblique to shaft & appears to be condylar [JS88]; ulna olecranon robust & expanded proximally [JS88]; ulna sigmoid & mediolaterally narrow (typical mammal) [JS88]; ulna shaft anteroposteriorly broad [JS88]; both epipodials with flat distal articulations & no styloid process [JS88]; pelvic acetabular dorsal emargination closed [H+97] (contra, [J+02]) (contra [JS88], dorsal margin "broadly open 'V'"); pelvic elements unfused within acetabulum [J+02]; no femoral neck, femoral head oriented dorsally [J+02] (contra [JS88] femoral head set off by flat, broad neck); lesser trochanter present as broad flange with unique deep fossa dorsally [JS88]; additional flange-like trochanter distal to greater trochanter [JS88]; broad, shallow patellar groove flanked by crest-like epicondyles [JS88]; fibula does not contact distal femur [J+02] (compare [JS88] proximal fibula "probably" contacted tibia, not femur); external foot "spur" as in Zhangheotherium and monotremes [H+97] [L+02, 1] [JS88]; sesamoid bone present in the pedal flexor tendon [H+97] (contra [J+02]); metapodials & phalanges short & robust [JS88].
Notes: [1] According to The Age of Dinosaurs Lives On, Gobiconodon was the largest Mesozoic mammal at 5 kg and 50 cm. [2] This odd character state is of obvious interest in finding the place of triconodonts. Unfortunately, the ulnar articulation is known only for a few taxa. Morganucodontidae, Cimolodonta derived Multituberculata), and the platypus Ornithorhynchus have the primitive condyle. The hybrid state is found in triconodonts, the symmetrodont Zhangheotherium and a few transitional forms (Vincelestes, Henkelotherium). Even more interesting, a similar transitional state is found for the radial articulation only in Zhangheotherium and the two transitional mammals. Triconodonts, monotremes and multituberculates have the primitive condylar arrangement for the radius. [J+02]. [3] There seems to be some uncertainty here. [K+00] mention no second incisor and state that both "i1 and c are semi-procumbent." This would suggest that Ji et al.'s i2 is Kielan-Jaworowska et al.'s canine. However, both sources identify 4 premolars. In fact [K+00] state that some specimens have only three. The explanation is provided by [JS88]. These authors believe that the first apparently premolariform tooth is actually a reduced canine. The caniniform tooth is a modified incisor. In addition, they assert that Gobiconodon had deciduous P4/p4 (P5/p5, if you count the premolariform canines) which was not replaced in adults, leaving a diastema between P3/p3 and M1/m1. These two peculiarities, plus the existence of replacement molars and the gradual (?) addition of molars distally during ontogeny, have created gross inconsistencies in descriptions of Gobiconodon's dentition. [4] This character should set off alarms. The "wrapping cingulum" character is almost precisely the same as the purported synapomorphy that [L+02] use to assert that the ausktribosphenids have an independently derived tribosphenic molar. See discussion at Australosphenida and related essay.
Links: Mesozoic Mammals; Docodonta and Triconodonta, an internet directory; Life Through Time Content; CENTRE DE RECHERCHES LORRAINES; 416165a-s1.doc; mammif chinois.pdf (pdf file of Hu et al., 1997); 416816a-s1.pdf; The Age of Dinosaurs Lives On.
References: Hu et al. 1997) [H+97]; Jenkins & Schaff (1988) [JS88]; Ji et al. (1999) [J+99]; Ji et al. (2002) [J+02]; Kielan-Jaworowska et al. 2000) [K+00]; Luo et al. (2002) [L+02]; Sigogneau-Russell (1995) [S95]. APW 040806
 Amphilestes: A. broderipii Owen 1859.
Amphilestes: A. broderipii Owen 1859.
Range: Middle Jurassic (Bathonian) of Europe (England).
Phylogeny: Triconodonta:: (Jeholodens + Triconodontidae) + *.
Characters: Meckelian groove present, converging with ventral border of the dentary anteriorly [C+99] [L+02]; coronoid process inclined at 125° [L+02]; dentary condyle massive & bulbous, broad in dorsal view [L+02]; 3-4 lower incisors, with last columnar [CM98]; upper canine unknown, lower canine, present, large, double-rooted [L+02]; 4 lower premolars [L+02]; last lower premolar with b much smaller than c [L+02]; last lower premolar with c distinct, but much shorter than a based on valley between them [L+02]; cusp a of ultimate lower premolar symmetrical [L+02]; labial cingulid absent or vestigial in ultimate lower premolar [L+02]; lower molars with labial face generally concave [F79]; lower molar a cusp dominates crown [S95]; cusps b&c displaced lingually [F79]; lower molar cusps a>b~c>d [S95]; molar a cusp occludes in groove between A & B [L+02]; height of c <40% height of a [L+02]; lower molars with mesiolingual cingular cuspule e [L+02]; lower molars with distinct lingual cingulid [L+02]; lower molars interlock, with d/hypoconulid fitting between e & f cuspules of next more distal molar [L+02]; carnivore
Links: Mesozoic Mammals; Triconodonta, an internet directory; Amphilestidae;
References: Cifelli & Madsden (1998) [CM98]; Cifelli et al. 1999) [C+99]; Freeman (1979) [F79]; Luo et al. 2002) [L+02]; Sigogneau-Russell 1995) [S95]. APW 040806
 Jeholodens: J. jenkinsi. Ji et al. (1999).
Jeholodens: J. jenkinsi. Ji et al. (1999).
Range: Early Cretaceous (Barremian) of China (Yixian Fm., Liaoning Province)
Phylogeny: Triconodonta::: Triconodontidae + *.
~12 cm; orbits large; sagittal crest present but not prominent [L+02, 1]; braincase bulges at level of parietals [L+02, 1]; dentary coronoid process inclined at 145° [L+02]; dentary condyle mediolaterally narrow, but deep & long [L+02]; i1 procumbent & enlarged [L+02]; small upper and lower canines present [L+02]; 2 premolars [L+02]; diastema between P1 & P2 [L+02]; main cusp a of ultimate lower premolar not symmetrical [L+02]; last lower premolar with b at least as large as c [L+02]; last lower premolar with c distinct, but much shorter than a based on valley between them [L+02]; labial cingulid absent or vestigial in ultimate lower premolar [L+02]; molar a cusp occludes in groove between A & B [L+02]; upper molars with A cusp larger than B&C, which are subequal [PM02]; M1 without labial or lingual cingulum [PM02]; M2 & M3 with faint buccal cingulum [PM02]; upper molars with distal cingular cusp D [PM02]; lower molar cusps b&c with long axis angled from axis of cusp a [PM02]; lower molars with posterior lingual cingulid of the lower molar absent or weak (reversal?) [L+02]; a cusp larger than b&c, which are subequal [PM02]; height of c <40% height of a [L+02]; lower molars with cuspules e & f absent [PM02]; lingual cingulid discontinuous [PM02]; lower molars interlock, with d/hypoconulid fitting into groove or embayment in cusp b of next more distal molar [L+02]; axis-atlas complex very primitive, including (a) atlas neural arch and intercentrum not fused, (b) atlas & axis ribs present, (c) prezygapophysis on atlas, (d) dens not fused to atlas, (e) post-axis cervical ribs present [L+02]; with 15+ thoracic vertebrae [L+02]; lumbar ribs not fused to vertebrae [L+02]; tail long; interclavicle present [L+02]; interclavicle overlaps manubrium ventrally but is not fused with it (primitively no overlap, fused in living mammals) [L+02]; interclavicle with median anterior process [L+02]; supraspinous fossa fully developed along entire dorsal border of scapula [L+02]; acromion process hook-like & extending below glenoid [L+02]; procoracoid fused to sternum & absent as separate element [L+02]; coracoid small, without posterior process [L+02]; glenoid articular surface at angle to scapula & faces mostly posterior (not laterally) [L+02]; glenoid rounded (not elongate) & concave (not saddle-shaped) [L+02]; forelimbs generally advanced; humeral head weakly inflected [L+02]; humeral shaft with strong torsion (>30°) [L+02]; humerus deltopectoral crest extending past midpoint of shaft [L+02]; humerus ulnar trochlea present posteriorly, with vestigial condyle anteriorly [L+02]; humerus epicondyles weakly developed [L+02]; "grasping" hands; hindlimbs generally sprawling & primitive; pelvic elements unfused [L+02]; femoral head without neck, oriented dorsally [L+02]; astragalar neck & trochlea absent [L+01]; calcaneal tubercle ("heel") short & without terminal swelling [L+02]; calcaneum anteromedial face contacts cuboid derived!) [L+02]; calcaneum sustentacular facet vertical & on medial edge [L+02, 0]; cuboid offset from MtV [L+02, 0]; Mt III points laterally compared to axis of calcaneum (as in image of Kryptobataar) (L+02, 1]; tarsal spur absent [L+2, 0]; insectivore.
Links: Paleontology and Geology Glossary: J; Roots of the Mammalian Family Tree; Nature Science Update; News in Science 25/3/1999 Ancient Chinese Mini-mammal; Mesozoic Mammals; Docodonta and Triconodonta, an internet directory; EXN Science Wire: Daily news from the world of science.
References: Luo et al. 2002) [L+02]; Prasad & Manhas (2002) [PM02]. APW 040806
{kind=link}
{kind=link}