 1. Inside the Jaw
1. Inside the Jaw| Mammaliformes | ||
| The Vertebrates | Symmetrodonta |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Cynodontia
│
└─Mammaliformes
├─Allotheria
│ ├─Haramiyida
│ └─Multituberculata
│ ├─Paulchoffatiidae
│ └─┬─Gondwanatheria
│ └─Cimolodonta
└─┬─Morganucodontidae
└─┬─Docodonta
│ ├─Megazostrodontidae
│ └─Docodontidae
└─┬─Hadrocodium
└─Symmetrodonta
├─Kuehneotheriidae
│
└─Mammalia
|
Contents
Overview |
Luo Zhe-Xi is the master of mammaliforms. [1] His papers over the last few years have transformed our understanding of the key transitions in the mid-Mesozoic which made us the hairy niusances we are today. Jeholodens, Zhangheotherium, and now Hadrocodium have all been described by Dr. Luo and his associates at the IVPP in Beijing and the Carnegie Museum in Pittsburgh. Hu et al. (1997); Qiang et al. (1999); Luo et al. (2001). There are, however, two trivial matters about his scientific output that are somewhat difficult to deal with. The first is that Dr. Luo is from the PRC. Consequently, his name (along with other researchers from China) has been Westernized in a number of different ways. As it turns out, LZ Xi, L Zhexi, Z-X Luo, and so forth are all the same Dr. Luo, although each scientific persona has probably published enough important papers to satisfy most workers.
The other matter has less to do with persons than economics. Luo publishes some of his best material in Science. This is a Good Thing, since more people see it this way. However, the page limitations in Science are severe. When Luo's Zhangheotherium paper came out, there was a certain amount of professional grumbling that the authors had actually revealed very little, particularly about the all-important teeth. Possibly in reaction, the Hadrocodium paper almost goes to an opposite extreme. It pushes the envelope on length at 5½ pages. More importantly, in an effort to shoe-horn the maximum amount of data per column-centimeter, the paper is more densely written than an annotated edition of Joyce's Ulysses. The cryptically abbreviated labels on the figures are in so fine a font that one has to enlist neighborhood children with good eyesight to decipher them. In addition, the paper comes with an on-line addendum of about 100 pages, although this is unfigured and largely concerns the cladistic analysis. For most readers of these Notes, it would be best to set aside a full day -- with a 200 watt bulb and a good magnifying glass -- to tackle this work.
Nevertheless, Luo et al. (2001) is a fine paper, or really two fine papers. The first part is the description of Hadrocodium -- an isolated skull -- and its mammalian adaptations. The second part is the mandatory cladogram. Both are worth careful study. However, for the purposes of this Note, we will restrict our attention to the first section. The object here is simply to unpack this part of the article a little and make the incredibly dense Figures 1 and 2 a bit more accessible.
Hadrocodium is tiny by any measure. It is fully adult but Luo et al. estimate the mass of the creature at 2g -- as small as the very smallest modern mammals. This may be significant. If one is looking for the source of major morphological innovations, one might expect to see more of them in a small organism with a very high rate of reproduction. Each individual represents a trivial investment of gestation time and food energy from the standpoint of the species, and genetic drift should be higher in species with a) short generation times and (b) populations which are easily separated by physical barriers. Thus selection against mutation may be relaxed, and those that occur are more likely to become entrenched in some meaningful number of individuals. The implication is that Hadrocodium may not simply be the sister of all mammals, but something very close to an actual ancestral form.
Luo et al. describe Hadrocodium as having four suites of mammalian apomorphies. The first two are quite simple. The third and fourth are interrelated and quite complex.
1. Inside the Jaw
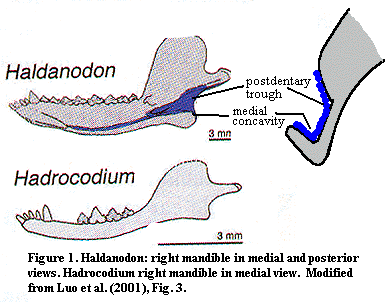
This series of changes is fairly straightforward. As we will discuss at great length below, the original postdentary bones in the ancestral synapsids were the angular, surangular and articular. The jaw joint was formed by an articulation between the articular and the quadrate. Between the Carboniferous pelycosaurs and the first Jurassic mammals, the jaw articulation changed. These post-dentary bones became increasingly small and less relevant to prey capture and eating. By the Middle Jurassic, Docodonts, such as Haldanodon, relied exclusively on a "new" dentary-squamosal jaw joint.
However, the lower jaw was also involved in sound reception, and it was advantageous to have small bones in this area. Through a series of transformations which need not be detailed here, the dentary overgrew the old postdentary elements. These, in turn, formed a small trough and concavity at the posterior end of the jaw, on its medial (inside) surface. This space supported the tympanic membrane on the tympanum (angular) as well as the auditory ossicles of the middle ear: the incus (quadrate), malleus (articular) and stapes.
Crown group mammals have all lost this portion of the jaw, and the entire assemblage has been transferred to the cranium. The key observation here is that Hadrocodium has no trough or concavity. The conclusion is that the middle ear machinery had already been moved to the cranium. As we will see, this supposition is supported by other anatomical observations. Interestingly, the dentary retains an odd, inflected curve that both mammals and other mammaliforms lack. The implication is that the loss of the postdentary elements from the jaw had perhaps been very recent, so that the dentary retained the odd curvature which would be left by simply subtracting out the portion formerly related to the postdentary bones. [2].
 2. The Brain
2. The Brain
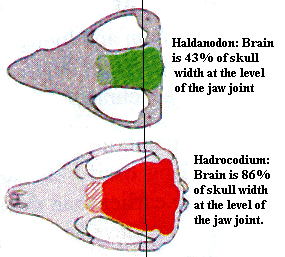
Mammal brains are bigger. This is one that everyone knows. Many steps were involved in this development, but the step represented by Hadrocodium seems to have been a large one. In mammaliforms below the level of Hadrocodium, the occiput, the jaw joint, and the transverse part of the zygomatic arch were all at more or less the same level of the skull. The squamosal portion of the arch flared out widely to accomodate the bulging jaw musculature. However, the muscles driving the jaw still took up a great deal of room.
In Hadrocodium, jaw articulation is moved anteriorly, so that the jaw musculature is spread forward as well, leaving more room for the brain cavity in the posterior half of the skull. The reason for this is very likely the separation of of hearing from eating. The jaw was able to move forward only because it was freed from its connection with the otic capsule. Even more important for later mammals, it became possible to expand the brain cavity posteriorly and dorsally, since the otic capsules could be moved away from the jaw articulation. We can see that this process has already begun in Hadrocodium. In addition, it is likely that the existence of the tympanum on the lower jaw imposed constraints on the muscle attachments to the jaw. Although we cannot know what these constraints were, it is certain that they were removed by the shift of the tympanum to the cranium.
3. The Jaw Joint and the Mammalian Ear
 Here's where things get a bit complex. We must begin with a long, digressive journey. As with many complex journeys in osteology, we start by paying a visit to our friend Bob, the Basal Amniote. Bob, as usual, has the Standard Condition. Looking closely at the lower corner of the back of Bob's head, we see that the quadrate is a key player. It braces the stapes, which supports the braincase through the otic capsule. Most importantly for our purposes, it forms the cranial part of the craniomandibular joint -- the jaw joint. It acts as a kind of spool, and the articular bone, at the very back of the lower jaw, hooks around it. All this tension and torsion is stabilized by the squamosal (which attaches to the occipital bones and the skull table further up) and by the the quadratojugal (which is anchored on the jugal and maxilla).
Here's where things get a bit complex. We must begin with a long, digressive journey. As with many complex journeys in osteology, we start by paying a visit to our friend Bob, the Basal Amniote. Bob, as usual, has the Standard Condition. Looking closely at the lower corner of the back of Bob's head, we see that the quadrate is a key player. It braces the stapes, which supports the braincase through the otic capsule. Most importantly for our purposes, it forms the cranial part of the craniomandibular joint -- the jaw joint. It acts as a kind of spool, and the articular bone, at the very back of the lower jaw, hooks around it. All this tension and torsion is stabilized by the squamosal (which attaches to the occipital bones and the skull table further up) and by the the quadratojugal (which is anchored on the jugal and maxilla).
This is a fine arrangement for blockheaded basal amniotes. However, when evolution started knocking holes in the skull, the squamosal and quadratojugal became less dependable supports. At the same time, the formerly robust stapes evolved into a more gracile bone devoted to hearing. These developments, in the Sauria, both allowed and required the quadrate to become more massive and self-supporting. Eventually, as in lizards and dinosaurs, the quadrates took over from the squamosals and became pillars of the osteological community, graciously supporting the posterior corners of the skull. [3].
Among the synapsids, things took a very different turn. The skull table remains solid, as there is no upper temporal fenestra. However, squamosal is sternly barred from the skull table by the intervening supratemporal, and the back of the skull is frequently slanted. As a result, the posterior of the skull tends to be held together transversely by the occipital bones, rather than vertically.
The effect on the jaw attachment is dramatic. As shown in the Dimetrodon at right, the squamo sal has grown down almost over the quadrate and become a plate-like, lateral element. The jugal has undergone a similar transformation, while the quadratojugal has disappeared. In the lower jaw, the post-dentary elements angular, surangular, prearticular and articular) have rotated down to met the quadrate, while the dentary itself remains relatively straight -- as it must in order for the teeth to close simultaneously. The result is that the post-dentary bones have almost become sub-dentary bones. Finally, the angular has effectively been doubled, or folded (the "reflected lamina" of the angular), creating a sort of hollow tube below the jaw.
sal has grown down almost over the quadrate and become a plate-like, lateral element. The jugal has undergone a similar transformation, while the quadratojugal has disappeared. In the lower jaw, the post-dentary elements angular, surangular, prearticular and articular) have rotated down to met the quadrate, while the dentary itself remains relatively straight -- as it must in order for the teeth to close simultaneously. The result is that the post-dentary bones have almost become sub-dentary bones. Finally, the angular has effectively been doubled, or folded (the "reflected lamina" of the angular), creating a sort of hollow tube below the jaw.
There is not a great deal of solid information on what all this sculpturing in the lower jaw was supposed to accomplish. Much later, it becomes intimately connected with hearing. However, many textbooks notwithstanding, its a real stretch to imagine an ear drum the size of an Olympic gold medal on the lower jaw communicating vibrational information (how?!) with the stapes, which is hidden behind the quadrate. [4]. Any noise loud and/or low enough to cause this tympanum to vibrate in a coordinate fashion would probably be detectable by other means -- such as by observing landslides and seismic fissuring.
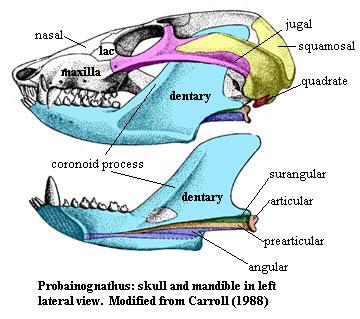 Whatever the answer, we must now press on about 50 My to the Middle Triassic and the eucynodont Probainognathus. Not by coincidence, Probainognathus is the outgroup used by Luo's group to root the phylogenetic analysis of Hadrocodium. At first glance, Probainognathus looks quite different from Dimetrodon. In reality, the changes are more matters of proportion than design. The temporal fenestra has become huge, dominating the greatly lengthened postorbital skull. The lateral skull is still composed of the squamosal and jugal, which bow outwards to form the zygomatic arch. In the lower jaw, the dentary has not only overgrown all of the other mandibular bones, but has formed its own, very large coronoid process. The coronoid bone has been reduced to a small, ovoid element on the medial face of the dentary. The splenial has vanished entirely. The dentary has also encroached on the postdentary bones ventrally, wrapping around them and leaving only long, thin remnants. These remnants include an angular, still with its miniature reflected lamina, forming a long concavity along the midline of the internal surface of the jaw. Despite the mass of the structure, the jaw still articulates through a small articular, which rotates on the quadrate, just as in Dimetrodon.
Whatever the answer, we must now press on about 50 My to the Middle Triassic and the eucynodont Probainognathus. Not by coincidence, Probainognathus is the outgroup used by Luo's group to root the phylogenetic analysis of Hadrocodium. At first glance, Probainognathus looks quite different from Dimetrodon. In reality, the changes are more matters of proportion than design. The temporal fenestra has become huge, dominating the greatly lengthened postorbital skull. The lateral skull is still composed of the squamosal and jugal, which bow outwards to form the zygomatic arch. In the lower jaw, the dentary has not only overgrown all of the other mandibular bones, but has formed its own, very large coronoid process. The coronoid bone has been reduced to a small, ovoid element on the medial face of the dentary. The splenial has vanished entirely. The dentary has also encroached on the postdentary bones ventrally, wrapping around them and leaving only long, thin remnants. These remnants include an angular, still with its miniature reflected lamina, forming a long concavity along the midline of the internal surface of the jaw. Despite the mass of the structure, the jaw still articulates through a small articular, which rotates on the quadrate, just as in Dimetrodon.
It seems unreasonable for a jaw of this size to rotate about so small an articulation. Probainognathus was not a large animal -- about 10 cm overall -- but the coronoid process is huge for the size of the jaw and the space available for the jaw adductors is enormous. To make things worse, the articular is not braced on the solid frame of the dentary, but on the feeble splinters of the surangular and prearticular. The system clearly worked, or these Notes would have been authored by something with scales and a muscular tail. But how it worked is a difficult matter. The engineering is usually explained by the decreasing importance of the jaw adductors and their gradual functional replacement by the masseter muscles, an evolutionary develoment which largely replaced slash-and-gulp eating with chewing -- so important to the development of civilized etiquette. Be that as it may, Probainognathus looks as if it posessed massive adductors and minimal table manners, so the issue is still incompletely resolved.
 Since we will be comparing Probainognathus to Hadrocodium, let us take a closer look -- first at the base of the skull and then at the jaw articulation itself. The quadrate and stapes are now both small elements. Although their size has changed, their relative position has not. The stapes clearly has no remaining significance in the structural support of the braincase. The stapes terminates at a well-defined fenestra ovalis, suggesting that the stapes was specialized for hearing. The otic capsule is otherwise relatively flat and featureless. The occiput has already fused to become a single ossification.
Since we will be comparing Probainognathus to Hadrocodium, let us take a closer look -- first at the base of the skull and then at the jaw articulation itself. The quadrate and stapes are now both small elements. Although their size has changed, their relative position has not. The stapes clearly has no remaining significance in the structural support of the braincase. The stapes terminates at a well-defined fenestra ovalis, suggesting that the stapes was specialized for hearing. The otic capsule is otherwise relatively flat and featureless. The occiput has already fused to become a single ossification.
With these landmarks in mind, examine a detail of the articular region. Two things should be apparant. First, when the articular contacts the quadrate, a corner of the dentary will contact the squamosal directly. Thus, even at the level of Probainognathus, we see the beginnings of the mammalian dentary-squamosal jaw joint.
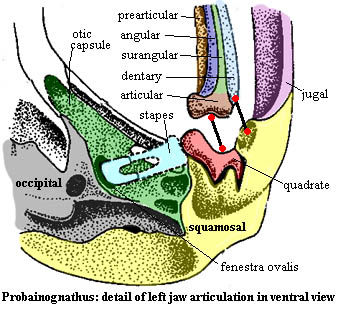 Second, the post-dentary bones are now inevitably going to have an impact on hearing. anything that shakes the articular will be carried through the quadrate to the stapes to the oval window and on to the inner ear. The thing most likely to set the articular vibrating is the long, thin surangular or its probable homologue in later forms, the retroarticular process of the articular. This stretches back (it wouldn't have far to go) to the point where the angular and its reflected lamina could now plausibly maintain a tympanic membrane of reasonable size.
Second, the post-dentary bones are now inevitably going to have an impact on hearing. anything that shakes the articular will be carried through the quadrate to the stapes to the oval window and on to the inner ear. The thing most likely to set the articular vibrating is the long, thin surangular or its probable homologue in later forms, the retroarticular process of the articular. This stretches back (it wouldn't have far to go) to the point where the angular and its reflected lamina could now plausibly maintain a tympanic membrane of reasonable size.
The rest, as they say, is history. The selection pressures which brought about the condition in Probainognathus are not really clear and took some 50 My to operate -- suggesting that these pressures were not particularly strong. However Probainognathus is only a few steps from the mammalian jaw and ear. The advantages of separate, specialized structures for chewing and hearing do not require much explanation. In fact, it is a little surprising that the mammalian condition took as long as 20-30 My more to develop. Perhaps this was due to a combination of two factors: (a) the rather more urgent matter of surviving the rise of dinosaurs in the Late Triassic and (b) the complex topological transformation needed to free the post-dentary bones from the dentary.
 Now, at last, we are ready to return to Hadrocodium. The changes from Probainognathus to Hadrocodium are more subtle, but in many ways more fundamental than the changes between Dimetrodon and Probainognathus. The postdentary bones have disappeared from the jaw without a trace. The mandible is now composed entirely of the dentary. The dentary articulates only with the squamosal through a fully developed glenoid. The quadrate is no longer in contact with the jaw, or even visible.
Now, at last, we are ready to return to Hadrocodium. The changes from Probainognathus to Hadrocodium are more subtle, but in many ways more fundamental than the changes between Dimetrodon and Probainognathus. The postdentary bones have disappeared from the jaw without a trace. The mandible is now composed entirely of the dentary. The dentary articulates only with the squamosal through a fully developed glenoid. The quadrate is no longer in contact with the jaw, or even visible.
All of these missing postdentary elements have presumably been incorporated into a very modern looking ear complex. The old otic capsule has been replaced by a much more complex structure, the petrosal, formed by fusion of the prootic, opisthotic, and incorporation of the postdentary elements. The paroccipital process has wrapped around the petrosal as a containing crista prootica. The tympanum is, of course, not preserved. However, the epitympanic recess is easily recognizable. Within this recess we would find the curved remnants of the angular (the tympanic) which actually form the tympanic annulus. The retroarticular process of the articular persists only as the tiny manubrium which remains in contact with the tympanic membrane as it presumably did in Probainognathus. The ancient articular - quadrate - stapes link is likewise preserved, but as the malleus - incus - stapes chain of ossicles in the mammalian middle ear. The entire assembly has been completely divorced from the jaw and is contained within the petrosal. Significantly, the inner ear has also expanded greatly and is easily observed as a large bulge, the promontorium. Finally, the promontorium bears a distinctive conical process for the cochlea, the signature organ of hearing in mammals.
This set of developments is really what sets Hadrocodium and mammals apart from all that came before. The separation of the jaw and ear freed the posterior skull from a significant restraint. Since the otic capsule was no longer under selective pressure to remain in exact coordination with the craniomandibular joint, the skull was free to expand posteriorly and laterally. The jaw, in turn, was freed to adopt configurations suited to take maximum advantage of precise dental articulations. That is, it was the separation of hearing and chewing that indirectly permitted the explosion of dental specializations which characterize evolution among the crown group mammals. --ATW 010616
Notes: [1] Luo refers to the taxon as "Mammaliaformes." This is probably incorrect. The members of Mammaliformes are not morphologically similar to Mammalia, which is a clade. They are morphologically similar to mammals. Thus, it is appropriate to use the combining form for the critters themselves, rather than their taxon. [2] The absence of the groove and re-engineering of the lower jaw should also result in repositioning the Meckelian cartilage. As we have seen elsewhere, this can result in novelties in dentition and jaw function which might be related to the evolution of the tribosphenic molar. [3] See, for example, the discussion at Hadrosauridae or the sumptuous illustrations in Fairman 1999). [4] At this point, the hopelessly compulsive may wish to review where this is all going by visiting The Ear.
References: Fairman 1999); Hu et al. (1997); Luo et al. (2001); Qiang et al. (1999); Wang et al. 2001).
lwJ of China.
Mammaliformes:::: Symmetrodonta + *.
Post-dentary foramen absent; post-dentary bones completely disassociated from jaw; fully functional temporomandibular joint; zygomatic arch does oriented anteriorly (does not swing laterally); cranial process of squamosal anterior to fenestra vestibuli; post -glenoid depression on squamosal between cranial & zygomatic processes; large promontorium (housing of cochlea) present on petrosal; epitympanic recess (attachment for incus) present; brain expanded laterally to mammalian proportions.
Image: Hadrocodium wui. Modified from a reconstruction by Mark Klinger. Length without tail: about 3 cm. Changes similar to those made to Morganucodon: outline leaner, slight crest added, shape of rostrum changed, pinnae reshaped and moved, vibrissae further de-emphasized. Possibly, the eyes could be more lateral. Klinger's original appears on the cover of Science for May 25, 2001. Since he is with the Carnegie and had the opportunity to work with Luo, his work is more accurate. The point here is simply to show an alternative.
Notes: Wang et al. (2001) regard Hadrocodium as a probable juvenile. ATW 010610.
Links:Flash Hadrocodium (Greek); Tiny Animal May Be Mammals' Ancestor.
References: Fairman 1999); Hu et al. (1997); Luo et al. (2001); Qiang et al. (1999); Wang et al. 2001).
Symmetrodonta: = Holotheria? = Kuehneotherium + Prof. Kuehn) Kotatherium family i.s.)
Range: upT-upK.
Mammaliformes:::: Hadrocodium + *: Kuehneotheriidae + Mammalia.
Tiny, very unspecialized mammals; cranial: auditory meatus may be present; cochlea primitively uncoiled; no angular process; loss or reduction of Meckelian groove and coronoid bone in derived forms; jaw action with some transverse and proplinal elements (?); primitively post-dentary bones in slot (?!!), but advanced forms converge on mammalian condition; $ molars with basic reversed triangles pattern [L+01]; molars fit into embrasures of teeth in opposite jaw; occlusal surfaces formed developmentally, rather than by wear; molars with two roots; cristae may bear complex pattern of cuspules; external (i.e. buccal) cingulum well-developed and may bear cusps [SE98]; upper teeth: buccal cingulum frequently absent [SE98]; upper molars with tall, lingual A/paracone [SE98]; mesiobuccal B/stylocone [SE98]; one or more buccodistal cusps including C/metacone, with or without a more distal 'c' conule [SE98]; lower teeth: $ buccal curvature of lower molar more bulging on primary a/protoconid than on cusps b/paraconid and c/metaconid [L+01]; $ lingual curvature of a/protoconid is far less convex than on b/paraconid c/metaconid [L+01]; $ hypoconid present [L+01]; may have small talonid, but talonid has probably been derived independently in different lines [L+01], is anterior in one case, and may or may not be homologous with mammalian (some derived forms have no talonid); $ posterior talonid is incipient, or with a cingulid or cingulid cuspule [L+01]; $ b/paraconid c/metaconid support single wear facet [L+01]; $ lingual cingulid present, at least distally [L+01] [SE98]; lower molar roots of equal size [SE98]; axial: cervical vertebrae not fused; thoracic ribs robust and antero-posteriorly compressed; appendicular: interclavicle primitively present, but allows clavicle to pivot shoulder blade; forelimb posture sprawling, but clavicles move broadly and independently; epipubics present; pelvic girdle slight & not deep; acetabulum with dorsal ridge; ankle spurs as in monotremes.
Links: Cifelli & Madsen; Sigogneau-Russell; Transitional Vertebrate Fossils FAQ: Part 1B; Evol.Page to Print: Clado Key: Team Assignments and Traits; Mammal origins; mammif_chinois.pdf; SyrenCRL French -- useful despite the lack of the figures it describes).
Discussion: To judge by the literature, symmetrodonts were a group of vaguely triangular, free-range molars that roamed the world unencumbered by either bodies or synapomorphies for almost 150My. In fact, until 1997 there was not a single symmetrodont bone known other than the occasional partial dentary. Many symmetrodont species exist only as tiny, isolated molars and, except for Zhangheotherium, the entire world's production of symmetrodont remains could probably fit into a beer mug.
To this day there is no satisfactory understanding of what a symmetrodont is. Much of the early work was done on Kuehneotherium. This genus was founded on a number of teeth and lower jaws found in association with fissure fillings of the basal mammal Morganucodon, whose teeth are frankly not very different. It was therefore supposed that Kuehneotherium looked pretty much like Morganucodon. The eventual discovery of virtually complete remains from the advanced, Early Cretaceous Zhangheotherium (Hu et al. (1997)) suggests that this wild guess was not far off. Certainly, Zhangheotherium has many interesting features, some of which are startlingly advanced. However, it looks pretty much like every other Mesozoic mammal.
Worse, as Sigogneau-Russell & Ensom (1998) note, there are no real and unequivocal synapomorphies which allow us to say what is symmetrodont and what isn't. Again, all symmetrodont species consist entirely of teeth and the odd dentary fragment, except for Zhangheotherium. For the latter, the teeth have yet to be described in meaningful detail. By definition, symmetrodont molars have the ancestral tricuspid configuration. The homologies between this molar type and the tribosphenic molar of Theria are not understood, and the nomenclature and description of these teeth is confused and inconsistent. For example, Fox's 1985) taxonomy seems [3] to refer to the primary cusp of the lower molars as both "paraconid" and "protoconid" on the same page. Because of these difficulties, as well as the persistent mammalian problem of convergences, Sigogneau-Russell and Ensom were unable to perform a cladistic analysis. It remains entirely possible that we are dealing, not with a discrete phylogenetic category, but with a series of intermediates between triconodonts, an the one hand, and dryolestoids and therians, on the other.
As a result, some phylogenies have essentially abandoned the whole idea of the symmetrodonts as a separate branch of the mammalian radiation. Instead, the various symmetrodont families are arranged as separate branches, or even paraphyletic grades, around the main trunk of mammalian evolution. There is nothing inherently wrong with such an arrangement. It might also help to explain some biogeographical anomalies in which similar new dental species show up more or less simultaneously on opposite shores of the Tethys and Labrador Seas. See, e.g., Sigogneau-Russell & Ensom (1998) (Tethys); Cifelli & Madsen (Labrador). Unfortunately, in most cases, treating the symmetrodonts as paraphyletic only makes matters worse. Land bridges may be a better explanation. However, Luo et al. (2001a) present powerful evidence that the symmetrodonts are simply the stem lineage of all mammals, and that the tribosphenic molar evolved twice (at least!) in this group.
Truthfully, there is not enough evidence for fully informed consent to either course at the moment. Since we do not know what species are correctly described as symmetrodont, it is difficult to make a rational choice. There does seem to be a core group of forms which originally developed a talonid of sorts, but subsequently minimized and lost this dental tool. Instead, they evolved in the direction of complex and poorly understood stylar interconnections between adjacent teeth, with the elaboration of stylar cuspules and ridges. These elements probably worked together to create very precise articulations between molars and the embrasures between teeth on the opposite jaw. It is also noteworthy that the incisors of symmetrodonts, where they are known, tend to show substantial specialization. One might speculate that, like rodents, the advanced symmetrodonts, particularly Spalacotheroidea, had at least two possible types of articulation depending on what set of teeth were being used.
However, this is all supposition. For the time being, symmetrodonts will be treated here as the stem lineage of mammalian evolution which diverged from other mammaliforms sometime in the Late Triassic or Early Jurassic. When Zhangheotherium is fully described, and, with luck, something other than a tooth is found in North America or Europe, we may see a very different picture. ATW 001230. Revised 011221.
Note: [1] a third, African, radiation of Symmetrodonts may also be present in the Late Mesozoic. [2] For dental terminology in symmetrodonts, see images at Kuehneotheriidae and Spalacotheroidea. [3] I am unable to rule out the possibility that I completely misunderstand what Fox is saying.
References: Ensom & Sigogneau-Russell (2000); Fox 1985); Hu et al. (1997); Luo et al. (2001a) [L+01]; Sigogneau-Russell & Ensom (1998) [SE98]. 020218.
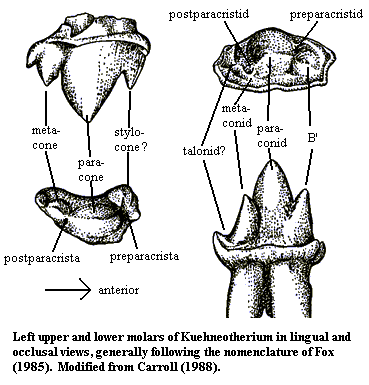 Kuehneotheriidae: "Obtuse-angle symmetrodonts." Kuehneotherium, Woutersia??
Kuehneotheriidae: "Obtuse-angle symmetrodonts." Kuehneotherium, Woutersia??
mT-lwJ of Eur.
Symmetrodonta: Mammalia + *.
Retain some trough for post-dentary bones [L+02]; both upper and lower molars have well-developed trigon and molar is triangular in cross-section, with three roots; angle formed by three principal molar cusps is generally >90°, with angle decreasing on posterior molars; paracone highest and most lingual on uppers; complete cingulum present [SE98]; cingulum without cuspules [SE98]; lower molars with a/protoconid (sometimes referred to as paraconid) largest and most buccal; small talonid present [SE98]; metaconid small (questioned by [SE98]); postparacristid shorter and more transverse than preparacristid; strong cingulids lingually & mesially, otherwise weak or absent [SE98]; believed to have had some degree of transverse jaw movement.
Links: mammalogy1.html; augtwoe; MESOZOIC MAMMALS; KUEHNEOTHERIIDAE and Co & "AMPHIDONTIDAE" Best on the Web); Mammals very basic); SyrenCRL; French); volume_2_3-4; Untitled (brief explanation of traditional take on this taxon).
References: Fox (1985); Luo et al. (2002) [L+02]; Sigogneau-Russell & Ensom (1998) [SE98]. ATW030101.
checked ATW050521
