Mammaliformes
Allotheria
Taxa on this Page
- Allotheria X
- Cimolodonta X
- Gondwanatheria X
- Haramiyida X
- Mammaliformes
- Multituberculata X
- Paulchoffatiidae X
Multituberculates
Multituberculates are like that enormous 20-piece fondue set that Aunt Harriet gave you as a wedding present. Its very pretty and well made, but it doesn't go with anything and there's really no place to put it. Pieces of it get used for other things, and keep being lost and found, but no one knows how to assemble it -- and when was the last time you had everyone over for fondue, anyway?
At first glance, multis look like mammals. The skull structure is relatively familiar. They have differentiated teeth which occlude and have familiar-looking cusps. they have a one-piece dentary jaw, a normal mammalian middle ear, zygomatic arch, and are the right size and shape. However, the appearance is deceptive. Miao (1993) shows dramatically how few characters multituburculates share with mammals. In fact, he finds only two: absence of the septomaxilla and presence of the three otic ossicles. The molars -- if that's really what they are -- are nothing like the molars of mammals. In fact, multis didn't even chew the same way. There is essentially no transverse, grinding movement involved. That type of motion derives from unilateral occlusion, where only one side of the jaw occludes at a time. Multuberculates always made contact on both sides of the jaw. Multis combined orthal chomping with a peculiar backstroke in which the cusps of the lower jaw were dragged posteriorly through the basins or along the sides of the upper teeth. Wall & Krause (1992). The multituberculate rostrum is also distinctly unmammalian. It is a box-like construction, more like a pelycosaur than a mammal, with the large flat maxillae forming the sides, the nasal the top, the the tall premaxilla at the rostral end. Compare the image of Dimetrodon. The zygomatic arch looks right, but the anterior portion is mostly maxilla, rather than jugal. The jugal is a minor element stuck in a little slot in the maxillary process. The squamosal never gets involved in the braincase, and the whole arrangement of the petrosal area is quite different from mammals.
 The post-cranial skeleton is not very well known, but no multi is known which even approached a parasagital gait -- and the foot has been radically redesigned for an even more sprawling posture than a protomammal. Imagine a furry little defensive lineman or a mouse with the posture of a sumo wrestler. Perhaps this is a bit misleading. Kielan-Jaworowska has speculated that some multis may have been great jumpers, and this is easy to believe. It depends on the overall build, and most multis were small and gracile, so far as we know. It is only the posture that reminds us of something big and blocky. Multis are often called the "rodents of the Mesozoic," but it might be more apt to think of them as the "rabbits of the Mesozoic."
The post-cranial skeleton is not very well known, but no multi is known which even approached a parasagital gait -- and the foot has been radically redesigned for an even more sprawling posture than a protomammal. Imagine a furry little defensive lineman or a mouse with the posture of a sumo wrestler. Perhaps this is a bit misleading. Kielan-Jaworowska has speculated that some multis may have been great jumpers, and this is easy to believe. It depends on the overall build, and most multis were small and gracile, so far as we know. It is only the posture that reminds us of something big and blocky. Multis are often called the "rodents of the Mesozoic," but it might be more apt to think of them as the "rabbits of the Mesozoic."
With all their strangeness, it would seem natural to put the multituberculates outside the mammals. However, despite every indication to the contrary, it is generally taught that they are just some lost tribe of mammal Isrealites with funny teeth. The problem here seems to be historical. As long ago as 1880, Marsh suggested a link between the multituberculates and "Microcleptes," later known as Haramiya, and erected the Allotheria to contain them. [1]. The haramiyids have a very spotty record. However, they do (mostly) have two parallel rows of cusps on their molariform teeth. Unfortunately, until very recently, that is all they had. Haramiya consisted only of a few teeth, as did every other genus of haramiyid. Thus, the argument for a particular pre-mammalian origin rested only on a few grams of tiny teeth, mostly from a single Late Triassic site in France. That wasn't too bad when the multis were just a tooth order as well, but the fossil record of multis kept improving, while the record of haramiyids did not. Thus, the connection between multituberculates and haramiyids was one of those issues like the dino-bird connection before the Chinese feather factory went into production: much debated, but with insufficient real evidence on either side.
To add to the difficulty, the definition of "mammal" is not generally agreed. The tendency is still to use a non-cladistic definition involving the posession of various "mammalian" character states. See for example, Pough et al. (1999). The discussion by those authors is excellent, but the fact remains that a non-cladistic definition invites problems with polyphyly. [2]. That is the reason that the definition of Mammalia in these Notes has recently been changed to include only crown group mammals, i.e. the last common ancestor of platypuses, possums & people and all of its descendants. Thus, under some definitions of "mammal", the multis would be mammals regardless of the haramiyid connection. However, that does not mean that they are more closely related to us than are echidnas.
In any case, the whole tree was shaken in 1997 with the description of Haramiyavia by Jenkins et al. (1997). Unlike every other haramiyid, Haramiyavia had jaws, and even bits and pieces of post-cranial skeleton. At last, haramiyid students had more than teeth to work on. Jenkins reconstructed Haramiyavia as having only an orthal jaw motion -- slash and gulp -- and other dental detail which appeared to be inconsistant with a multituberculate connection. "Available evidence," they concluded, "does not indicate a close relationship between the two groups."
This would appear to have killed off the Allotheria. However, the Allotheria still have a champion in Percy Butler, professor emeritus of the University of London, nor was he slow to answer from the barge. Professor Butler must be very nearly the oldest of the Old Guard, as he is now [July, 2001] almost 90. Although he claims that "I know no geology, and have never collected a fossil," he nonetheless managed to collect the Romer-Simpson Medal, the highest honor granted by the Society for Vertebrate Paleontology, back when he was a mere youth of age 82.
Prof. Butler may not know geology, but he certainly knows fossil teeth, which he has studied for some 65 years. In particular, he was one of the first to look at mammaliform teeth as products of ecological adaptation and developmental biology. Butler (2000) carefully compared the teeth of Haramiyavia and Thomasia. Neither Jenkins nor anyone else disputes that these forms are closely related. In addition, Butler was able to include data from a new haramiyid, Eleutherodon, which had not been available to Jenkins. Butler determined what the homologies were between the cusps in the various haramiyids. Using this information and some rather detailed dental modelling, Butler shows that there is a reasonable progression from orthal chomping, with a small degree of palinal motion in Haramiyavia to the fully palinal motion in multituberculates.
Of course, having a reasonable story to tell is not the same as evidence, but it does seem to disprove Jenkins' assertion that there is no possible relationship between haramiyids and multis. What is missing is a cladistic analysis. One suspects that it may not be possible to do this yet. As discussed elsewhere, it is critically important to get a good handle on the homologies before doing this analysis. Butler goes far in this direction, but we still lack enough bones from enough taxa to tell the difference between a reasonable conjecture and good working hypothesis.
In the mean time, the Allotheria remain the best guess about the relationships involved. Jenkins' identification of a groove in the jaw for the post-dentary bones confirms the suspicion that the haramyids cannot be mammals. All three living groups of mammals have the post-dentary elements fully incorporated in the middle ear. As Luo et al. (2001) have elegantly shown, this incorporation predates the crown group. Thus, just as Miao (1993) suggested, the superficial similarities between multituberculates and mammals are probably convergences [3]. ATW 010708
Note: Zhelestids were the first placentals to specialize in herbivory and may have have dominated the small herbivore niche in the upK of Central Asia, a semitropical lowland and coastal environment. At the same time, multituberculates occupied the same niche in the more arid, upland environment of Mongolia. [K+00]. [1] Marsh, OC (1880), Notice on Jurassic mammals representing two new orders, Am. J. Sci. 3 20: 235-239, as stated in Butler 2000). The work of Jenkins et al. (1997) confirmed suspicions that two haramiyids, Haramiya and Thomasia, were actually the upper and lower teeth of the same animal. Since Thomasia has seniority, there is no Haramiya any more -- although there are still haramiyids. [2] This seems to be an odd trap that even the various best scientists fall into. Consider for example, the Arctometatarsalia of Holtz (1995) or the Myopterygia of Janvier. For an explanation of the problem, see Holtz (1996a). [3] Convergent development in this area may be particularly likely for reasons discussed elsewhere.
Descriptions
Mammaliformes: Sinocodon + living monotremes + living therians. [L+02].
Phylogeny: Probainognathia: Tritheledontidae + *: Allotheria + (Morganucodontidae + (Docodonta + (Hadrocodium + Symmetrodonta))).
Primitively, very small (<100g); $ fully developed dentary-squamosal jaw joint [L+02]; $ petrosal promontorium develops at expense of ventrolateral wing of basisphenoid [L+02]; diphyodont teeth; $ teeth double-rooted, with precise occlusion permits true chewing, hence very fine oral food processing); $ each post-canine tooth occludes with two teeth in opposite jaw; differentiated molars and pre-molars; fixed dental formula; prismatic enamel (very durable); dentary-squamosal jaw joint dominates; stapes very small; parietals fused; postorbital and prefrontal absent; brain enlarged; otic capsule (prootic + opisthotic) fused as petrosal; cerebellum folded; optic lobes divided; ring-shaped atlas; epiphyses on long bones; heart completely divided into four chambers; single aortic trunk; muscular diaphragm present; hair; sebaceous and/or sweat glands; endothermic (some exceptions known); typically viviparous; $ lactation and suckling; primitively nocturnal insectivores.
Links: The Hall of Mammals; Animal Diversity Web: Class Mammalia; Mammal Directory; Skulls Unlimited International Inc.; Mammals - Introduction; The Structure and Classification of the Mammalia (1864); General Zoology - Subphylum Vertebrata, Class Mammalia (very basic); Taxonomy browser (Mammalia); The Origin of Mammals.
It is in fact hard to draw the line; and, after you have finally drawn a clear line, some inconsiderate field scientist will immediately find something new and make a mess of your system. See, for example, the hints of weird things to come (out of Brazil) from Barbarena & Bonaparte: NAPC Abstracts, Ar - Be. Thus, both "Mammalia" and "Mammaliformes" (or "Mammaliaformes," although we deprecate that spelling) can and have been used in many different ways. Re- mammalian lineages (was Re- Cretaceous taeniodont). One of the problems with Mammaliformes is that some people, for various reasons, don't like the term. See Re- Cladospeak (Mammalia, Crurotarsi); and the glossary entries at Trevor Dykes' site. And, speaking of Trevor, probably the best coverage of the group (whose name he doesn't even like!) is at Trevor Dykes' MESOZOIC MAMMALS; Basal Mammaliaformes, Morganucodontidae ... page. Dr. Tom Holtz's notes, as usual, provide a good, short introduction to the characteristics of the group. Another good set of lecture notes (from Prof. Jon Baskin) can be found at Biology 4429 - Chapter 3. On specific organ systems, Dr. Luo Xhe-Zi provides a brief discussion of the evolution of the mammalian ear at CMNH Vertebrate Paleontology, while the text (no figures) of Christian Sidor's review of jaw changes can be found at Evolutionary trends and the origin of the mammalian lower jaw. Finally, Spotlight on Zoo Science- Lactating on Eggs - National Zoo| FONZ has an interesting, if a bit speculative, section on the evolution of lactation.
References: Luo et al. (2002) [L+02].
Note: Mammaliforms are unique among major taxa in that we have an enormous amount of information on one of the most basal species, Morganucodon. While this is obviously a Good Thing for scientists, it also makes it impossible to summarize "what makes a mammaliform" in a short paragraph. ATW050810.
Allotheria: Eleutherodon.
Molariform teeth with two rows of cusps arranged longitudinally; buccal row of lower molariforms bites bilaterally between rows of uppers; orthal and palinal chewing only, with little or no transverse, grinding element.
Links: Allotheria; SistEspEvol Portuguese); Main Page; Lecture 03 - Cont. Drift; Untitled Document; MESOZOIC MAMMALS; Haramiyida, an internet directory:; gatesy_pictures/11.JPEG;
References: Butler 2000), Miao (1993).
Note: supported by Miao basically by default as a place to put multis. 011202.
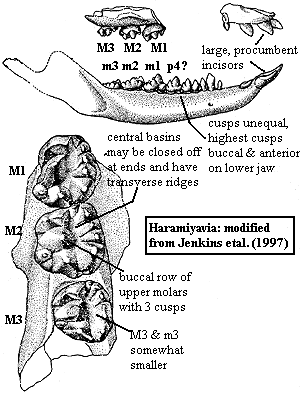 Haramiyida: Eleutherodon, Haramiya, Haramiyavia, Theroteinus, Thomasia.
Haramiyida: Eleutherodon, Haramiya, Haramiyavia, Theroteinus, Thomasia.
upT (Norian)-upJ of Eur, Greenland & Africa.
Procumbent incisors (3/1-3); cheek teeth with 2+ roots; crown broad with cusps along margin, frequently in two parallel rows enclosing a central basin; 1st 2 molars of top and bottom larger than 3rd; at least one lower molar has central basin, rather than central row cusps; basins tend to be closed posteriorly, open anteriorly; molariform cusps of unequal height, with largest cusps anterior buccal on lower row, posterior or median lingual on upper row; buccal row of upper molariforms with 3 cusps, with middle cusp tallest except Eleutherodon); occlusion pattern with cusps meeting opposing basins, usually the tallest upper lingual cusp in the basin of the lowers and/or the highest buccal lower in the basin of the uppers; jaw movement primarily or entirely orthal; dentary gracile & arcuate in Haramiyavia (looks a good deal like Hadrocodium, with coronoid a bit lower); sulcus for postdentary bones present; zygomatic arch originates above P3-P4 as in paulchoffatiids (actually "M3-M4" since these are all molariforms, rather than differentiated Ps & Ms); distal humerus with 2 condyles; well-developed olecranon process; gracile compared to Morganucodon, with longer jaw; humerus about same length as radius.
Links: Haramiyidae.
References: Butler 2000); Jenkins et al. 1997).
Note: May be mammaliform lineage unrelated to multituberculates or, alternatively, paraphyletic parent of multituberculates. Present majority view is probably that they are unrelated to multituberculates and represent an early mammaliform offshoot. The possibility exists that haramiyids are an early offshoot and that they are closely related to multituberculates. This is the position of Butler (2000) and is followed here. This approach does more to explain the weird primitiveness of multis. Per Jenkins et al. (1997), Thomasia is the lower dentition of Haramiya. 010701.
Multituberculata(= Plagiaulacoidea): Paulchoffatia + Cimolodonta.
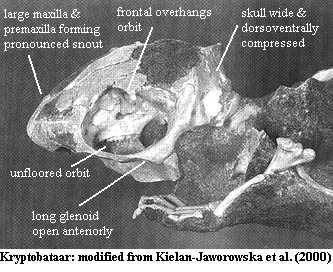 Range:upJ (mJ?)-lwOc of Eur, Asia, NAm, & SAm.
Range:upJ (mJ?)-lwOc of Eur, Asia, NAm, & SAm.
Mesozoic equivalent of rodents. Size 2-20 cm, up to several kg; herbivorous and omnivorous; skull low, broad & consolidated; large, strong jaw; angular process absent; no transverse jaw motion; dental formula 1-3/1, 0-1/0,1+/0+, 2+/1+; one pair of lower incisors enlarged & procumbent; opposite of rodents in that half-enameled lower incisors may exist independent of upper incisors (i.e. they may have gnawed with lower teeth!); mandibular symphysis not solid; no canines except upper in basal forms; large diastema; buccal cusps on lower premolars reduced or absent; large, blade-like premolars with multiple serrations; premolars clearly differentiated from molars; large multi-cusped molars with as many as three rows of cusps & 5-30 total cusps on a molar; cusps of roughly equal height; last upper molar is displaced lingually in relation to last lower; some dental specializations including blade-like forms; dentary "semicircular" and arcuate; jaw glenoid large, flat, tear-drop shaped & open  anteriorly (allows propalinal motion); mandible brought posteriorly into articulation; choanae separated by vertical plate formed by vomers; most of posterior palate formed by palatines; ectopterygoids fused with pterygoid?; septomaxilla absent; $ premaxilla large; maxilla large and forms most of lateral and ventral snout; maxilla usually with one infraorbital foramen; $ nasal and nasal cavity large (not long) in most; nasal cavity with complex ventral ridge system (turbinates?); cribiform plate absent?; frontal overhangs orbit; palatine excluded from orbit (i.e., orbital process of palatine absent); orbits face laterally; orbits probably of moderate size and unfloored; orbits extended anteriorly; $ postorbital process reduced or absent; postorbital process on parietal; $ jugal reduced & zygomatic arch largely from squamosal and maxilla (jugal is internal to arch); squamosal small & contributes little to braincase; zygomatic arch stout, with jugal incorporated in medial wall; parietal extensive on skull roof but not significant otherwise; post-glenoid region short and wide; olfactory lobes large, with large, tall petrosal; supraglenoid foramen located on raised lateral wall of petrosal; 3 auditory ossicles as in therians; stapes not perforated and "columellaform"; stapes with round footplate; bone bulla absent; no bone external auditory meatus; promontorium narrow, straight & oriented anteromedially; cochlea nearly straight; vestibular apparatus differs radically in size among multis and may be greatly inflated; internal auditory meatus very small or absent; alisphenoid small (controversial); cerebrum smooth; no cerebellar hemispheres; cochlea uncoiled & ear relatively primitive; vertebrae with wide transverse and high spinous processes thus jumper??); interclavicle present; scapula lacks supraspinous fossa; glenoid large, opening ventrally; pelvis very narrow (too narrow for shelled egg per Kielan-Jawarowska); "marsupial" bones present; acetabulum shallow & dorsal (from Ptilodus: may be arboreal adaptation rather than plesiomorphic); parafibula present in knee area as in monotremes; calcaneum contacts MtV; MtIII abducted ~30° from longitudinal axis (i.e. deeply sprawling posture); wide variety of ecological niches including arboreal form (Ptilodus) with reversed hallux and possible prehensile tail; also saltatorial and fossorial forms; possibly viviparous based on pelvic morphology. Longest lived mammalian lineage.
anteriorly (allows propalinal motion); mandible brought posteriorly into articulation; choanae separated by vertical plate formed by vomers; most of posterior palate formed by palatines; ectopterygoids fused with pterygoid?; septomaxilla absent; $ premaxilla large; maxilla large and forms most of lateral and ventral snout; maxilla usually with one infraorbital foramen; $ nasal and nasal cavity large (not long) in most; nasal cavity with complex ventral ridge system (turbinates?); cribiform plate absent?; frontal overhangs orbit; palatine excluded from orbit (i.e., orbital process of palatine absent); orbits face laterally; orbits probably of moderate size and unfloored; orbits extended anteriorly; $ postorbital process reduced or absent; postorbital process on parietal; $ jugal reduced & zygomatic arch largely from squamosal and maxilla (jugal is internal to arch); squamosal small & contributes little to braincase; zygomatic arch stout, with jugal incorporated in medial wall; parietal extensive on skull roof but not significant otherwise; post-glenoid region short and wide; olfactory lobes large, with large, tall petrosal; supraglenoid foramen located on raised lateral wall of petrosal; 3 auditory ossicles as in therians; stapes not perforated and "columellaform"; stapes with round footplate; bone bulla absent; no bone external auditory meatus; promontorium narrow, straight & oriented anteromedially; cochlea nearly straight; vestibular apparatus differs radically in size among multis and may be greatly inflated; internal auditory meatus very small or absent; alisphenoid small (controversial); cerebrum smooth; no cerebellar hemispheres; cochlea uncoiled & ear relatively primitive; vertebrae with wide transverse and high spinous processes thus jumper??); interclavicle present; scapula lacks supraspinous fossa; glenoid large, opening ventrally; pelvis very narrow (too narrow for shelled egg per Kielan-Jawarowska); "marsupial" bones present; acetabulum shallow & dorsal (from Ptilodus: may be arboreal adaptation rather than plesiomorphic); parafibula present in knee area as in monotremes; calcaneum contacts MtV; MtIII abducted ~30° from longitudinal axis (i.e. deeply sprawling posture); wide variety of ecological niches including arboreal form (Ptilodus) with reversed hallux and possible prehensile tail; also saltatorial and fossorial forms; possibly viviparous based on pelvic morphology. Longest lived mammalian lineage.
Links: Introduction to Multituberculates; pal1; mammevol.htm; kryptobataar; digital reslicing of kryptobataar; The first furry creatures; Main Page; Mesozoic mammals showcase; Literature - Mammalia; UTDMG
- Kryptobaatar dashzevegi; Paleocene mammals of the world (Best on the Web); Stars of the Show (A to I); Untitled Document.
References: Butler 2000), Jenkins et al. 1997); Holtz (1995); Holtz 1996a); Kielan-Jaworowska et al. (2000); Luo et al. (2001); Miao (1993); Pough et al. (1999); Simmons (1993); Wall & Krause (1992). ATW 011128.
Paulchoffatiidae: sensu Simmons (1993), i.e. Paulchoffatia only.
upJ (Kimmeridgian) of Eur (Guimarota).
Dental formula 3/1, 1?/0, 5/3, 2/3; enamel evenly coats incisors;I3 on margin of palate, with multiple cusps; C1 present; P2-P4 double-rooted; P2 with 5-6 cusps; p3 & p4 have 3-5 cusps; p4 & m1 very short; p4 rectangular; lower premolars (p1-p3) not fully differentiated for shearing; M2 short, without lingual cusps; at least 1 molar (here $ m2) with central basin instead of linear cusps (like haramiyids, so quite possibly plesiomorphic); incisive foramina small & round or oval; palatal vacuities absent; angle of coronoid >55° from tooth row; maxilla has two infraorbital foramina; no bone process (of frontal) over orbit; frontals pointed anteriorly & rounded posteriorly (?), postorbital process reduced or absent; nasal relatively compressed laterally; lacrimal present, large & extends to frontals; frontal-parietal suture V-shaped; zygomatic ridge weak or absent; zygomatic arch flares out & is not confluent with snout; base of zygomatic arch dorsal to P3-P4 embrasure; $ nasal contacts parietal; $? condylar foramina (canal for XIIth [hypoglossal] nerve) paired .
Links:Plagiaulacoidea; MESOZOIC MAMMALS; Plagiaulacidae and Paulchoffatiidae, an internet directory:; SyrenCRL French).
References: Kielan-Jawarowska & Hurum (1997); Simmons (1993).
Note: "Plagiaulacoidea" is clearly paraphyletic as confirmed by virtually all studies. As a result that taxon is not used here. Paulchoffatiids are the most basal known multis. 020228.
Gondwanatheria: Ferugliotherium, Gondwanatherium, Lavanify, Sudamerica.
upK-lwPc of SAm, India, Mad & probably Antarctica.
large, gnawing incisors; many with hypsodont dentition; characteristic microstructure of molar enamel, involving small circular prisms separated by sheets of matrix; molars curved along their height; molars with deep infundibulum; transverse ridges between molar cusps; p3 absent?; rectangular p4.
Links: scienceweek; Paleocene mammals of the world (Best on the Web); Untitled Document questioning whether actually related to multis).
References: Kielan-Jaworowska & Hurum (1997); Krause et al. (1997). 010701.
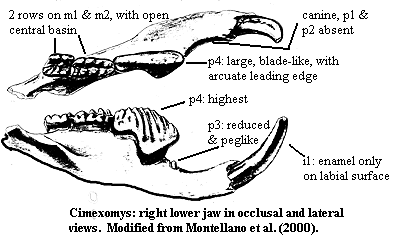 Cimolodonta: Cimexomys, Ptilodus.
Cimolodonta: Cimexomys, Ptilodus.
upJ?-lwOc (mainly upK-upPc) of China & NAm.
$? I1 absent; $ enamel on i1 only on labial surface (?! -- see Wall & Krause (1992) on Ptilodus); $? I2 with <4 cusps; $ I3 located medially;$ no canines; $ P0 absent; $ p2 absent; p3 small & peg-like (Cimexomys); $ p4 has arcuate leading edge; $? p4 monophyodont; $? M2 labial cusp row present; substantial incisive foramen in premaxilla; $ palatal vacuities present; $ snout length >50% of skull length; $ 1 pair of infraorbital foramina; prenasal process of premaxilla forms internarial bar; $? lacrimal absent; $? frontal forms roof over anterior of orbit; $ jugal fossa (on zygomatic arch?) large and deep; $ strong zygomatic ridge; low condyle relative to occlusal plane maximized force at p4 (highest lower tooth) for crushing & slicing; probable omnivores with some degree of specialization for medium-large food objects; lacrimal present in one early group; vestibular apparatus may be enlarged; $ single pair of condylar foramina; vertebrae dorso-ventrally compressed (Cimexomys).
Links: Cimolodonta.
References: Kielan-Jaworowska et al. (2000); Kielan-Jawarowska & Hurum (1997); Miao (1993); Montellano et al. (2000); Simmons 1993); Wall & Krause (1992).
Notes: [1] This clade is, obviously, very well supported. [2] The Djadochtatheria, Mongolian upK multis, were originally proposed to fall outside Cimolodonta. Kielan-Jawarowska & Hurum 1997). However, their cladogram actually places Djadochtatheria within Cimolodonta, and the group has accordingly been moved to this position. Kielan-Jaworowska et al. (2000). [3] In this general connection, the humble editor of these Notes would like to express his undying gratitude to Drs. Kielan-Jawarowska and Sigogneau-Russell for never co-authoring a paper. The world of early mammalian paleontology has been immeasurably enriched by the efforts of each scholar individually, but the small world of scientific commentary is made easier by not having to pronounce both formidable names in the same breath. [4] Wall & Krause (1992) analyze the jaw cycle in Ptilodus as: (a) jaw opened and moved forward on the glenoid (~2.5 mm); (b) orthal movement to either (i) pierce with incisors or ii) crush and slice with the elaborate multituberculate p4; (c) retraction of the jaw with grinding along molar surfaces. Osteology suggests that posterior temporalis muscle was very strong and primarily responsible for adduction, masseter & temporalis for retraction. 010703.
{kind=link}