 Morganucodontidae: Brachyzostrodon, Morganucodon (= Eozostrodon?)
Morganucodontidae: Brachyzostrodon, Morganucodon (= Eozostrodon?)| Mammaliformes | ||
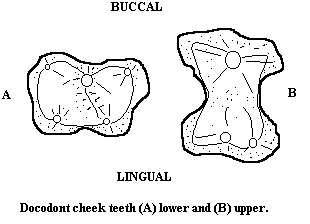
| The Vertebrates | Docodonta |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Cynodontia
│
└─Mammaliformes
├─Allotheria
│ ├─Haramiyida
│ └─Multituberculata
│ ├─Paulchoffatiidae
│ └─┬─Gondwanatheria
│ └─Cimolodonta
└─┬─Morganucodontidae
└─┬─Docodonta
│ ├─Megazostrodontidae
│ └─Docodontidae
└─┬─Hadrocodium
└─Symmetrodonta
├─Kuehneotheriidae
│
└─Mammalia
|
Contents
Overview |
Morganucodontidae: Brachyzostrodon, Morganucodon (= Eozostrodon?)
Range: upT-mJ of Eur, China, Russia & NAm(?)
Phylogeny: Mammaliaformes:: Docodonta + (Hadrocodium + Symmetrodonta)) + *.
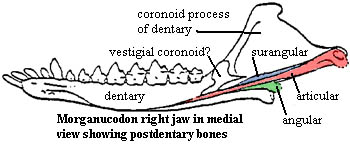
Characters: General small, rat-like forms, ~10cm long; skull <3 cm, long & slender; Dentition dental formula 5/4, 1/1, 4/4, 4/4; well-differentiated, diphyodont molars; three cusps on the upper and lower molars; cusps were aligned anteroposteriorly with precise occlusion; 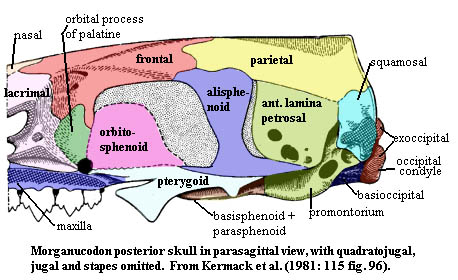 Jaws slender jaw; dentary loosely attached at symphysis; post-dentary bones present in groove of dentary; post-dentary bones ~20% of jaw length (?); coronoid bone present at base of coronoid process of dentary; reflected lamina decreased to narrow ribbon-like horseshoe; both quadrate-articular and squamosal-dentary jaw joints; possible tympanic membrane in reflected lamina of angular; Palate hard palate from fused maxillae & fused palatines only; posterior palate shows ridges as in tritylodonts & multis; strong pterygoid flanges as in cynodonts limited lateral motion in reptilian jaw); ectopterygoids may be present primitive) no gap between palate & basicranium (change from cynodonts); Snout nares confluent (?); septomaxilla unknown but believed present; large nasal cavity above secondary palate, with ridges for turbinals (so clearly warm-blooded); narrow snout; probable vibrissae (from pattern of nerve & vessel foramina); maxilla large and tall (as multis); large nasal with fine grooves for circulation, suggesting fleshy snout; lacrimal large, forming anterior root of zygoma with maxilla; Orbit orbits large; no postorbital bar; temporal fenestra merged with eye socket; postorbital and prefrontal absent; anterior and
Jaws slender jaw; dentary loosely attached at symphysis; post-dentary bones present in groove of dentary; post-dentary bones ~20% of jaw length (?); coronoid bone present at base of coronoid process of dentary; reflected lamina decreased to narrow ribbon-like horseshoe; both quadrate-articular and squamosal-dentary jaw joints; possible tympanic membrane in reflected lamina of angular; Palate hard palate from fused maxillae & fused palatines only; posterior palate shows ridges as in tritylodonts & multis; strong pterygoid flanges as in cynodonts limited lateral motion in reptilian jaw); ectopterygoids may be present primitive) no gap between palate & basicranium (change from cynodonts); Snout nares confluent (?); septomaxilla unknown but believed present; large nasal cavity above secondary palate, with ridges for turbinals (so clearly warm-blooded); narrow snout; probable vibrissae (from pattern of nerve & vessel foramina); maxilla large and tall (as multis); large nasal with fine grooves for circulation, suggesting fleshy snout; lacrimal large, forming anterior root of zygoma with maxilla; Orbit orbits large; no postorbital bar; temporal fenestra merged with eye socket; postorbital and prefrontal absent; anterior and dorsal margins from lacrimal & frontal, respectively; anteromedial surface from ascending process of palatine; medial surface from partial orbitosphenoid wall; Skull table frontal large, with circulation grooves as on nasal, frontals meeting dorsal process of palatine; frontal and parietal extend ventrolaterally over dorsal braincase; parietals fused, forming sagittal crest; transverse (lambdoid) crest formed by posterior parietals; pineal foramen absent; squamosal does not participate in braincase (like multis, unlike therians); Lateral skull zygomatic arch slender and dorsally arched but not much expanded laterally; most of anterior arch (except root) from thin jugal; posterior arch from squamosal; Jaw articulation both dentary-squamosal and articular-quadrate articulations present; jaw articulation far posterior (as therians) with little post-zygomatic skull; Occiput strong lambdoid crest from parietal & dorsal flange of squamosal; foramen magnum large; basioccipital has little occipital exposure; most of foramen magnum margin and all of occipital condyles from exoccipitals; condyles low compared to therians; posttemporal fenestra reduced to minor opening between dorsal & quadrate flanges of squamosal; jugular foramen present between exoccipitals & petrosal; tabulars and supraoccipital unknown but probably present; Braincase cribiform plate apparently present; orbitosphenoid partially (?) ossified within orbit; no contact wit
dorsal margins from lacrimal & frontal, respectively; anteromedial surface from ascending process of palatine; medial surface from partial orbitosphenoid wall; Skull table frontal large, with circulation grooves as on nasal, frontals meeting dorsal process of palatine; frontal and parietal extend ventrolaterally over dorsal braincase; parietals fused, forming sagittal crest; transverse (lambdoid) crest formed by posterior parietals; pineal foramen absent; squamosal does not participate in braincase (like multis, unlike therians); Lateral skull zygomatic arch slender and dorsally arched but not much expanded laterally; most of anterior arch (except root) from thin jugal; posterior arch from squamosal; Jaw articulation both dentary-squamosal and articular-quadrate articulations present; jaw articulation far posterior (as therians) with little post-zygomatic skull; Occiput strong lambdoid crest from parietal & dorsal flange of squamosal; foramen magnum large; basioccipital has little occipital exposure; most of foramen magnum margin and all of occipital condyles from exoccipitals; condyles low compared to therians; posttemporal fenestra reduced to minor opening between dorsal & quadrate flanges of squamosal; jugular foramen present between exoccipitals & petrosal; tabulars and supraoccipital unknown but probably present; Braincase cribiform plate apparently present; orbitosphenoid partially (?) ossified within orbit; no contact wit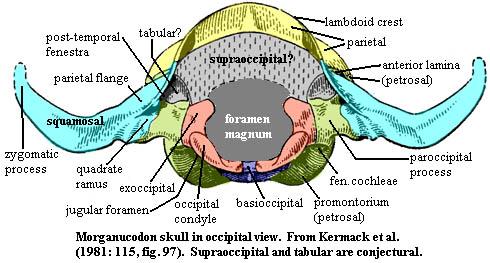 h ascending process of alisphenoid; narrow, strap-like alisphenoid; ascending processes of alisphenoid on either side of sella turcica; petrosal lies behind alisphenoid (see discussion for details); basicranium triangular (primitive), dominated by basioccipital; lateral sides of triangle formed by strong basipterygoid processes, merging posteriorly with quadrate (posterior) rami of alisphenoid; internal corotid foramina on ventral surface of basisphenoid; basisphenoid lacks definitive dorsum sellae; basisphenoid sutured to pterygoids; parasphenoid (and vomer?) fused to pterygoid; lateral portion of basicranium from petrosal; enlarged cochlea with promontorium;
h ascending process of alisphenoid; narrow, strap-like alisphenoid; ascending processes of alisphenoid on either side of sella turcica; petrosal lies behind alisphenoid (see discussion for details); basicranium triangular (primitive), dominated by basioccipital; lateral sides of triangle formed by strong basipterygoid processes, merging posteriorly with quadrate (posterior) rami of alisphenoid; internal corotid foramina on ventral surface of basisphenoid; basisphenoid lacks definitive dorsum sellae; basisphenoid sutured to pterygoids; parasphenoid (and vomer?) fused to pterygoid; lateral portion of basicranium from petrosal; enlarged cochlea with promontorium; cochlea not coiled; auditory bulla absent (developed to confine auditory ossicles when separated from lower jaw); stapes extends from inner ear capsule to quadrate; stapes still large; quadrate tiny; petrosal with large anterior lamina; cerebral hemispheres small and smooth, not overlying cerebellum; Axial cervival vertebrae with very large neural canal; mammalian pattern of posteriorly slanted neural spines on anterior vertebrae, reverse in posterior vertebrae; clearly differentiated lumbar area without ribs; lumbar zygapophyses tilted 35-45° to resist torsion; ~27 presacral vertebrae; 2-3 sacrals; ~12 caudals; short tail; Appendicular limbs slender; procoracoid retained, but excluded from glenoid; no scapular spine; interclavicle and large clavicle retained; humerus broad, with ends twisted 50°; proximal head bent strongly dorsally; ectepicondylar foramen lost; entepicondylar foramen retained; pubes reduced; obturator foramen enlarged; short legs; femur essentially therian; 5-toed feet; Other probable fur (further extrapolation from vibrissae); insectivore(?).
cochlea not coiled; auditory bulla absent (developed to confine auditory ossicles when separated from lower jaw); stapes extends from inner ear capsule to quadrate; stapes still large; quadrate tiny; petrosal with large anterior lamina; cerebral hemispheres small and smooth, not overlying cerebellum; Axial cervival vertebrae with very large neural canal; mammalian pattern of posteriorly slanted neural spines on anterior vertebrae, reverse in posterior vertebrae; clearly differentiated lumbar area without ribs; lumbar zygapophyses tilted 35-45° to resist torsion; ~27 presacral vertebrae; 2-3 sacrals; ~12 caudals; short tail; Appendicular limbs slender; procoracoid retained, but excluded from glenoid; no scapular spine; interclavicle and large clavicle retained; humerus broad, with ends twisted 50°; proximal head bent strongly dorsally; ectepicondylar foramen lost; entepicondylar foramen retained; pubes reduced; obturator foramen enlarged; short legs; femur essentially therian; 5-toed feet; Other probable fur (further extrapolation from vibrissae); insectivore(?).
Links: Morganucodon Printout- EnchantedLearning.com; Morganucodon oehleri (Norwegian); Stark-Verlag German); Biological Principles, BI-120 Study Guide Page 20; Morganucodontidae after McKenna & Bell, 1997; Eozostrodon; Cuffey 2 - Mammal - Like Reptiles.
We have often blundered into braincase anatomy in these Notes, usually to back out again as quickly as our injured dignity will allow. However, we can advance no further into the issues peculiar to mammalian evolution without delving somewhat more deeply into these minutiae. Such practical considerations aside, it is also futile to spend all of our time charging full-tilt through phylospace, looking for the Big Pictures. It is the nature of phylospace that each Big Picture is made up of a very large number of Little Pictures, each of which is not only made up of yet smaller images, but connects with other issues altogether; so that what might appear to be an eternal answer at one scale, is merely one aspect of an infinite question at another. This being the case, it is appropriate to stop periodically to look carefully at some randomly chosen detail in order to gain a sense for the way in which it is tied to the rest of phylospace.
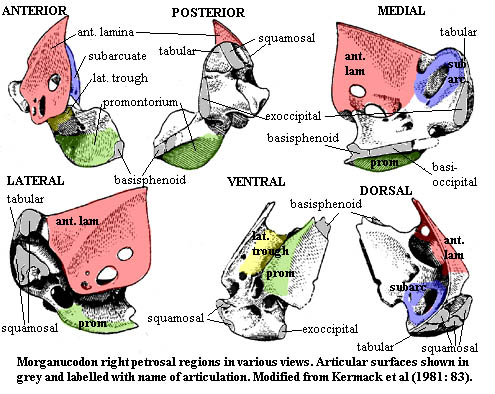 To be sure, the petrosal of Morganucodon could not fairly be described as "randomly chosen." Twenty years after the comprehensive work of Kermack et al. (1981), Morganucodon remains the world's best known mammaliform skull. The genus was originally known from one well-preserved and perfectly articulated skull. It was also discovered that various Paleozoic strata in Wales had been pierced by fissures in Early Jurassic times. These fissures had been filled by water-borne debris containing, among other things, hundreds of tiny, disarticulated bones from Morganucodon, many of which were in near-perfect condition. [1] The combination of a well-articulated and more or less complete skull and a broad sample of disarticulated bones allowed Kermack to reconstruct the head in unprecedented detail. Of all of the bones recovered by Kermack's group, the three, almost complete, petrosals are perhaps the most interesting for several reasons. First, the petrosal is the otic capsule: a fusion of (at least) the prootic and opisthotic bones. It is therefore the site of the crucial transfer of post-dentary bones from the jaw to the braincase to form the mammalian middle ear. This transfer had not yet occurred in Morganucodon; but it is supposed that the postdentary bones were already largely devoted to hearing. Second, the petrosal forms the lateral wall of most of the braincase, and thus reflects the path and position of many important cranial nerves and circulatory structures.
To be sure, the petrosal of Morganucodon could not fairly be described as "randomly chosen." Twenty years after the comprehensive work of Kermack et al. (1981), Morganucodon remains the world's best known mammaliform skull. The genus was originally known from one well-preserved and perfectly articulated skull. It was also discovered that various Paleozoic strata in Wales had been pierced by fissures in Early Jurassic times. These fissures had been filled by water-borne debris containing, among other things, hundreds of tiny, disarticulated bones from Morganucodon, many of which were in near-perfect condition. [1] The combination of a well-articulated and more or less complete skull and a broad sample of disarticulated bones allowed Kermack to reconstruct the head in unprecedented detail. Of all of the bones recovered by Kermack's group, the three, almost complete, petrosals are perhaps the most interesting for several reasons. First, the petrosal is the otic capsule: a fusion of (at least) the prootic and opisthotic bones. It is therefore the site of the crucial transfer of post-dentary bones from the jaw to the braincase to form the mammalian middle ear. This transfer had not yet occurred in Morganucodon; but it is supposed that the postdentary bones were already largely devoted to hearing. Second, the petrosal forms the lateral wall of most of the braincase, and thus reflects the path and position of many important cranial nerves and circulatory structures.
Study the first figure. If you are still searching for a Big Picture, this is it. The figure shows the petrosal in all of the usual orientations. Several arbitrary "regions" have been marked off as an aid to orientation and discussion, and the articular surfaces have been noted. For further assistance, you may wish to review the figure of the posterior skull in ventral view.
We might, to a very inaccurate first approximation, think of the petrosal as a sailing vessel, an old-fashioned gaff-rigged sloop, beating to windward against a stiff, posterolateral breeze, out through the occiput. The anterior lamina (in red) is the mainsail. The region of the subarcuate fossa (in blue) forms the foredeck and jib -- for the non-nautical, the little sail in front -- and the promontorium (green) marks the keel. The analogy breaks down when we try to describe the lateral trough (yellow), but, then, it wasn't much of an analogy in the first place.
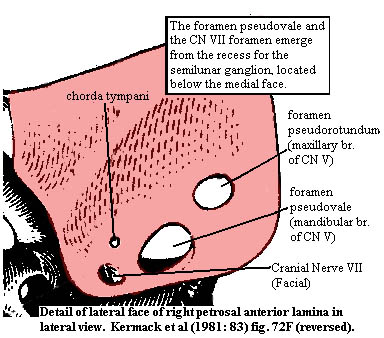 The anterior lamina
The anterior laminaThe anterior lamina (in red) might better be called the "lateral lamina." Its exact shape is a bit hard to make out from Kermack's figures. One is tempted to guess that it may be a crista prootica gone berserk. It seems to serve much the same function in sheltering the numerous neural fossae in the lateral trough, although it is much larger and more vertically oriented than a conventional crista, and it articulates with the parietal along its dorsal ridge (this articulation is not shown in the figure). The anterior limit of the anterior lamina must have approached the ascending process of the alisphenoid, but the two were not in sutural contact.
In lateral view, the anterior lamina is pierced by two large foramina: the foramen pseudorotundum anteriorly and the foramen pseudovalis posteriorly. Kermack notes that this identification has been disputed, but we will follow his system for lack of any basis on which to disagree. The two foramina are the exits of the maxillary and mandibular branches of the trigeminal nerve (cranial nerve V), respectively. The large pit at the base of the medial face of the anterior lamina housed the semilunar (or trigeminal) ganglion -- the nerve center from which the trigeminal nerves arise. The two smaller foramina on the anterior lamina accommodate the facial (VII) nerve and one of its branches, the chorda tympani, which carries taste sensations. The main facial nerve passes through the more ventral (lower) of the two small foramina and emerges in the pit for the semilunar ganglion.
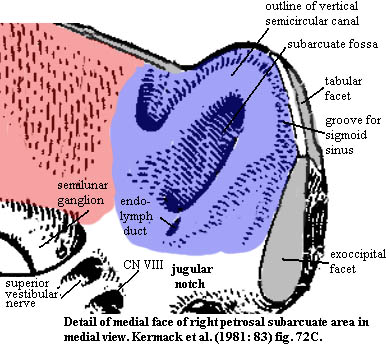 The subarcuate region
The subarcuate regionThe subarcuate region (in blue) has been colored only on its medial face. The central feature of this region is the subarcuate fossa, probably so called because it is the area under the arch of the vertical semicircular canal. The foramen for the chorda tympani on the lateral face of the anterior lamina emerges inside the subarcuate fossa, and, in life, the fossa was probably filled with sensory nerves from the chorda tymapni, carrying taste information from the mouth.
The small foramen just anteroventral to the subarcuate fossa is a branch of the endolymphatic system. Continuing in this direction, we find twin foramina which seem to approach obliquely from the anterior. The lower of the two is the auditory (VIIIth) nerve, and the upper is one of its branches, the superior vestibular nerve. The latter carries information about orientation and angular momentum from the lateral semicircular canal.
Immediately below (ventral) to the subarcuate fossa, the surface is somewhat recessed and the ventral edge of the petrosal is interrupted by a large, semi-circular notch, best seen in the dorsal view. This is the jugular notch, the petrosal half of the jugular foramen through which the internal jugular vein passes as it exits the cranium. The internal jugular then joins with the vena capitis lateralis [2] and drains into the neck to join the subclavian vein in the shoulder. The other half of the jugular foramen is formed by the exoccipital, the occipital, or the occipital portion of the temporal, depending on just how mammalian the organism is. The internal jugular drains the blood supply of the brain and originates from the sigmoid sinus. [3]. The jugular foramen also provides the exits for the glossopharyngeal (IXth) and vagus (Xth) cranial nerves. These mixed (i.e. motor and sensory) nerves are involved in brain interactions with the tongue and throat and with the viscera, respectively.
The lateral (uncolored) side of the subarcuate region is largely devoted to the articular surfaces for the squamosal and tabular. It projects a short way outward laterally. This lateral projection is all that remains of the opisthotic contribution to the paroccipital process in advanced mammaliforms. The exoccipital facet is perched on a small flange which extends a short ways out from the medial face. The entire articular region of the subarcuate area is fractured by vascular canals. The identity of the vessels is not completely clear. They certainly included the sigmoid sinus, which ran between the subarcuate fossa and the tabular facet, and the vena capitis lateralis, which may have been in the large canal almost directly opposite the subarcuate fossa on the lateral face.
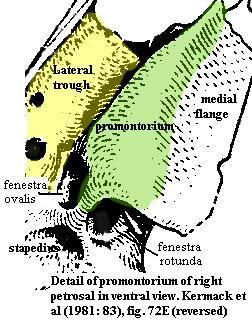 The promontorium area
The promontorium areaThe promontorium is the portion of the petrosal which houses the cochlea. The cochlea, a specialized and elongate process of the lagena, is the primary organ of hearing in mammals. In modern mammals, the cochlea is strongly coiled, a state which seems to be correlated with a sophisticated ability to discriminate among sounds. In mammaliforms, the cochlea is not coiled. The promontorium forms a sharp ridge anteriorly, but has a much more gradual curve toward the posterior end.
In ventral view, the promontorium ends posteriorly at the near-confluence of three large fenestrae: the fenestra rotunda, the fenestra vestibuli; and a pit which may have accommodated the body of the m. levator hyoidei. This muscle is a homologue of the mammalian stapedius muscle. Like the tensor tympani, the stapedius acts to damp excessively loud sounds -- in this case by restricting the movement of the stapes. The two middle ear fenestrate are discussed in connection with The Ear. Briefly, the footplate of the stapes fills the fenestra vestibuli and transmits sound to the inner ear. The fenestra rotundum simply moves in the opposite sense in order to relieve the pressure on the perilymphatic fluid in the inner ear caused by movement of the stapes.
Medial to the promontorium is a relatively featureless area, the medial flange. The edge of this flange provides the articular surfaces for the basisphenoid, anteriorly, and the basioccipital. Laterally, the promontorium forms the inner wall of the lateral trough.
Most of the foramina in the lateral trough cannot be identifies with the kind of certainty which applies to other petrosal features discussed here, nor does Kermack provide us with a definitive image of the area. According to these authors, the area is also somewhat variable. Accordingly, we will omit a detailed consideration of the lateral trough at this point.
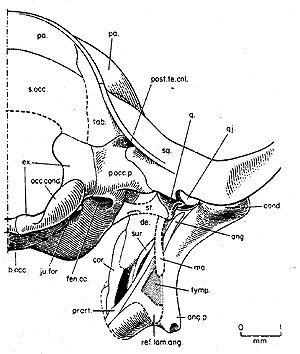 The beat of a different ear drum
The beat of a different ear drumThose who have been attempting to follow the ongoing struggle in these Notes to really understand braincase anatomy may be expecting that, at this point, we might finally have enough information to deal seriously with the development of the mammalian ear. That was, in fact, the original intent of this essay. Kermack et al. do provide their own vision of the pre-mammalian ear, involving a tympanum originally held by the angular and its reflected lamina distally, and by the manubrium (the retroarticular process of the articular), proximally. The difficulty is that the geometry and anatomy are simply not yet convincing.
The figure at right is that vision. Here are some of the problems. (1) Neither the reflected lamina of the angular nor the manubrium were fossilized, so that the entire critical structure is quite speculative. (2) The thing that is supposed to make the stapes vibrate appropriately is the quadrate. However, in Morganucodon, as in reptiles, the quadrate is still held in place by the quadratojugal which buffers it against stress with ligaments and soft tissues -- which would also seem certain to damp out sound vibrations imparted to the quadrate by the post-dentary bones. (3) As Kermack's group makes clear in another figure, the illustrated mechanism requires that at least the lower border of the tympanum be free. The physics of two-dimensional vibrating surfaces are quite complex [4]. However, no oscillator can operate with a free end or edge. How much music can you make with a guitar string if only one end is attached to the instrument? Essentially the same principle applies. (4) There is a reason why musical drums are round. Vibrations reaching the edge are reflected back and reinforce the vibrational node(s). The ear drum should work the same way. The trapezoidal to rectangular drum envisioned by Kermack can't work that way because the waves reflected from the edges will create a hash of rather randomly interfering signals. (5) Not to belabor the obvious, but is the space between the jaw and the skull really the best place to put an ear?
There may well be answers to these questions. For one thing, I am not certain that the tympanum really needs to have the properties of a harmonic ocillator, which would eliminate most of concerns (3), (4), and perhaps (2). Other answers are not obvious from our current level of understanding; nor do any alternatives immediately suggest themselves. Accordingly, we will once again have to leave this issue without a really satisfactory ending to the story. --ATW 011111
Notes: [1] The average cranial bone of Morganucodon is perhaps 2mm in maximum dimension. [2] That is, the "vein of the side of the head." The reader will not be stunned to learn that this vein drains the side of the head. [3] The sigmoid sinus itself resides in the mastoid process, a structure that will not be discussed here. [4] The reader may be aware that the famous Schrödinger wave functions are based on the equations describing standing waves in three-dimensional harmonic oscillators. For a graphic, see the link. The two-dimensional case is not really a heck of a lot simpler. See this link. I believe Scientific American ran an article on Chinese gongs about ten or fifteen years ago that touched on some of the math, but I lack the citation.
References: Kermack et al. (1981) [K+81]. 011102.
 Docodonta: Dinnetherium.
Docodonta: Dinnetherium.
mJ-upK of Eur, NAm & SAm.
Mammaliformes::: Hadrocodium + Symmetrodonta) + *: Megazostrodontidae + Docodontidae.
mouse-sized; snout & jaws elongate; probably retained some articular-quadrate contact with jaw; angular process of dentary directed ventrally; some species with as many as eight molars per jaw; lower molars rectangular and elongate; outer cusps taller than inner (derived from cingulum?), with ridge connecting buccal and lingual sides; upper molars expanded laterally with waisted central portion.
Links: Mesozoic Mammals; Docodonta and Triconodonta, an internet directory.
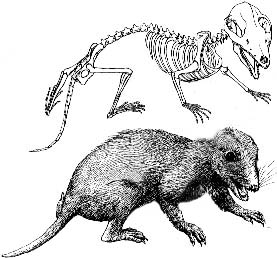 Megazostrodontidae: Megazostrodon.
Megazostrodontidae: Megazostrodon.
Range: upT to lwJ of Afr.
Phylogeny: Docodonta: Docodontidae + *.
Characters: ~10cm; dental formula 5/4,1/1,4/4,4/4; cheek teeth differentiated into premolars and molars; occlusal pattern in which upper teeth alternate with the lower teeth; dentary - squamosal functional jaw joint; articular & quadrate retained in jaw and skull base, respectively; brain with relatively large olfactory & auditory areas; relative brain size 3-4x cynodonts; and it had a flexible backbone with clear division into thoracic & lumbar regions; probable nocturnal insectivores.
Links: Paleontology and Geology Glossary: Me; Megazostrodon, Drawing Nature Studio; Megazostrodon; South African Museum - Fossil Reptiles of the South African Karoo; Evol.Page to Print: Clado Pic.Sinoconodon & Megazostrodon; first mammals appear; Biology 356; Mesozoic Mammals; Docodonta and Triconodonta, an internet directory; JURÁSSICO mamíferos (Spanish).
Image: Megazostrodon, substantially altered from Notes for Lecture 11. Auditory pinnae have been removed (can you imagine an external ear on the jawbone?), a small parietal crest added, the jaw gape is increased, the posture of all limbs made slightly more primitive, some superfluous shading removed, and body contour made leaner.
 Docodontidae: Docodon, Haldanodon, Reigitherium.
Docodontidae: Docodon, Haldanodon, Reigitherium.
Range: mJ-upJ of Eur. & NAm.
Phylogeny: Docodonta: Megazostrodontidae + *.
Characters: wear surfaces on mesiolingual part of lower molars, including basined crushing surface in some (reverse of normal mammal pattern in which talonid is posterior to to trigonid) [L+02]; omnivores(?).
Links: The Fossil Mammal Hall; Guimarota - A Jurassic Ecosystem (German); Pascual et al..
References: Luo et al. (2002) [L+02].
Image: Haldanodon, life reconstruction and left jaw in medial view. 020223.
checked ATW050521