 The Petrosal of Trioracodon
The Petrosal of Trioracodon| Mammalia | ||
| The Vertebrates | Triconodontidae |
| Vertebrates Home | Vertebrate | Vertebrate |
Abbreviated Dendrogram
Mammaliformes
│
└─Mammalia
├─Australosphenida
│ ├─Ausktribosphenidae
│ └─Monotremata
└─┬─Triconodonta
│
├─Jeholodens
│ └─Triconodontidae
│ ├─Priacodon
│ └─Trioracodon
└─┬─Spalacotheroidea
└─Cladotheria
├─Dryolestoidea
└─Theria
│
├─Metatheria
└─Eutheria |
Contents
Overview |
"Triconodontidae" has historically been used as a tooth taxon, including all triconodonts having molars with three roughly equal primary cusps. Triconodontid molars exhibit a strong trend toward elaboration of the accessory cuspules. These are not, as in tribosphenics, stretched out to form a protocone - trigonid mortar and pestle grinding system. Instead, the accessory cuspules are shaped to reinforce the interlocking system between adjacent molars. The central cusps still occlude with the spaces betweenadjacent opposite molars. The chewing mechanism is still primarily shear, and the cutting edges between cusps are formed by wear rather than being genetically determined cristae/cristids.
Richard Cifelli, with various co-authors, has argued that the Triconodontidae are polyphyletic. According to Cifelli, the Triconodontidae, as classically defined, minimally includes (a) a group possible clade) of Euro-American Jurassic triconodontids and (b) a separately derived clade of North American Cretaceous genera. However, the recent spate of new triconodont genera suggests that triconodont diversity is considerably undersampled. It may be too early to say what could and could not be derived from the Jurassic Triconodontidae. Furthermore, the current cladistic studies involving triconodontids are not reliable for the reasons discussed on the previous page. For these reasons, we will retain the conventional division for the present.
In fairness, we are not exactly in the best position to be making fatuous remarks about triconodontid ancestry and diversity. Our coverage of the triconodontids is presently limited to exactly two genera, Priacodon and Trioracodon, both of which are Jurassic. These are the only triconodontids with well-known non-dental remains. Of these, Priacodon has some unusual limb and girdle elements which were discussed on the previous page.
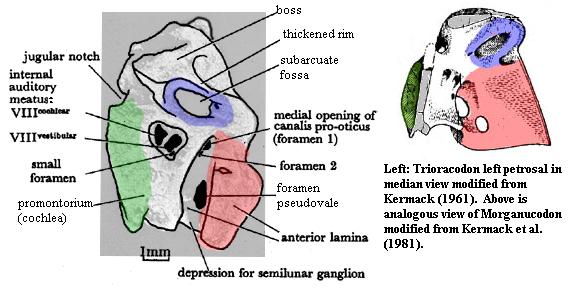
The Petrosal of TrioracodonLike the petrosal of Morganucodon, the petrosal of Trioracodon has been carefully described by Kermack 1963). Those readers who already bear the ritual scars of our mind-numbing struggle with the petrosal of Morganucodon will now receive their just reward. The Trioracodon petrosal differs only in detail. For those who have not yet undergone this essential rite of purification, turn back now before it is too late! "This quest is not for thee." [1]
Even more so than in Morganucodon, the anterior lamina of the petrosal is medially convex, leaving deep fossa for the semilunar ganglion, the root node of the trigeminal (Vth cranial) nerve. The internal auditory meatus is identical in form and location to its counterpart in Morganucodon, with ventral and dorsal foramina for the cochlear and vestibular branches of the auditory (VIIIth cranial) nerve, as well as a third foramen for an unknown blood vessel. The lateral face of the anterior lamina is pierced by a foramen pseudovale, exactly as in Morganucodon. However, foramen pseudorotundum is not clear in Trioracodon. As in Morganucodon, the petrosal has a large subarcuate fossa with a thickened rim of bone. By comparison with Morganucodon, the subarcuate fossa is somewhat smaller and has an anterodorsal slant. This may correlate with a life-style making fewer demands on the organs of balance.
On the other hand, the promontorium is larger and more complex. Hearing was perhaps a more significant part of Trioracodon's sensory repertoire. In the image, the promontorium appears to show some periodic changes in surface texture. These are not noted by Kermack, are not noticeable in the original photograph and may be artifacts of image preparation. However, there is some possibility that these markings are real, inadvertently enhanced by digital processing, and represent the beginnings of coiling or curvature in the cochlea. Interestingly, these markings appear to be restricted to the promontorium.
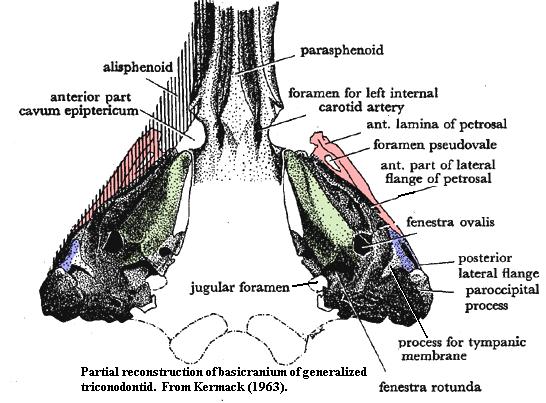
Kermack's notes, at some length, that the Trioracodon petrosal shows little qualitative change from the cynodont condition. This reasoning is based on the persistence of a cavum epiptericum. In cynodonts, neither the dermal bones of the skull nor the alisphenoid actually participate in the braincase. Posteriorly, the quadrate ramus of the cynodont alisphenoid runs along the inside wall of the dermal skull. It is braced, and separated from the braincase, by the lateral flange of the petrosal. More anteriorly, the outer margin of the braincase is formed by the pila antotica, a relict portion of the ancient sphenethmoid. The space between the internal braincase and the alisphenoid is the cavum epiptericum. The cavum contained, among other things, blood sinuses, and distal ganglia of the trigeminal nerve. In mammals, the brain is expanded. The outer wall of the braincase becomes the alisphenoid and the dermal skull bones. The structures of the cavum are incorporated into the brain. The lateral flange of the petrosal and the pila antotica are lost.
As in cynodonts and Morganucodon, the petrosal of Trioracodon retains a lateral flange (see image under Triconodontidae). It is small, and the posterior part of the cavum epiptericum appears to have been squeezed almost up against the alisphenoid. However, a noticeable lateral notch appears more anteriorly, and Kermack asserts that a pila antotica was present. The notch then represents the remaining space of the cavum. As Kermack convincingly notes, selection for even larger brain size leaves only two possibilities as the alisphenoid becomes convergent with the braincase. Either the anterior lamina continues to form the outer wall of the braincase, in which case the alisphenoid may be eliminated as redundant, or the alisphenoid becomes the new wall and the anterior lamina is abandoned. The former configuration was evolved by monotremes. The latter, by therians. [2]
Actually, we're not compelled to have one. Quite frequently we simply rattle on, in a self-indulgent manner, for absolutely no reason at all. Nonetheless, this is one of those occasions which does lend itself to a phylogenetic moral. That moral is as follows.
The phylogeny of Luo et al. 2002) has multituberculates, symmetrodonts, and triconodonts all emerging out of a chaotic region between Australosphenida and stem therians. That region is unstable, and involves an intuitively unreasonable mix of taxa. At least with regard to the Triconodonta, it does not seem to be supported by therian synapomorphies. We have looked in some detail at teeth, the forelimb, and the braincase. Except for some features of the scapulocoracoid apparatus, there is almost nothing which seems to require this phylogenetic result. And even there, some characters point towards monotremes rather than therians as the sister group of the triconodonts. The dental characters (92 of 275 total characters) are problematic. The 28 pectoral characters are ambiguous, and the whole design of the braincase is clearly primitive, as compared with either monotremes or therians. Most of the other characters involve huge amounts of missing data.
Triconodonts may be crown group mammals. We have, in fact, placed them among the mammals, for lack of a better place to put them. However, the evidence for this placement is still very weak. APW 040806
[1] "Thereafter, the dark warning of our King,
That most of us would follow wandering fires,
Came like a driving gloom across my mind.
Then every evil word I had spoken once,
And every evil thought I had thought of old,
And every evil deed I ever did,
Awoke and cried, 'This Quest is not for thee.'
--Tennyson, The Idylls of the King
[2] Lohnes et al. 1994) believe they have atavistically reconstructed the pila antotica, the cavum epiptericum, and the quadrate ramus of the alisphenoid in mouse embryos by genetic manipulation.
 Triconodontidae: = Triconodontinae). Alticonodon, Arundelconodon, Astroconodon, Corviconodon, Jugulator, Triconodon
Triconodontidae: = Triconodontinae). Alticonodon, Arundelconodon, Astroconodon, Corviconodon, Jugulator, Triconodon
Range: Middle Jurassic to Late Cretaceous of Europe (all Jurassic) & North America (mostly Cretaceous).
Phylogeny: Triconodonta::: Jeholodens + *: Priacodon + Trioracodon.
Characters: for braincase, see essay on Trioracodon & [K63]; Meckelian groove primitively present but lost independently (?) in several Cretaceous genera [R+01]; foramen at anteroventral margin of masseteric fossa [C+98]; dentition mono- or diphyodont, with replacement at very early age [R+01] [1]; i1 not specialized (i.e. not enlarged or procumbent) [L+02]; large upper and lower canines present [L+02]; primitively p4m5? [CM98]; premolars typically have mesial root much larger than distal [CM98]; main cusp a of ultimate lower premolar not symmetrical [L+02]; last lower premolar with b much smaller than c [L+02]; $ last lower premolar with c about as tall as a based on valley between them [L+02]; labial cingulid absent or vestigial in ultimate lower premolar [L+02]; main cusps of about equal height in both upper & lower molars [C+98]; buccal cingula -ids) reduced on both upper & lower molars [C+98]; molars added sequentially at distal end in some forms [R+01]; molars adult early in life [R+01]; molar a cusp occludes in groove between A & B [L+02]; upper molars with cuspule E/metastyle lost [L+02]; uppers interlock by tongue in groove mechanism [L+02]; lower molars with tongue-in-groove molar interlock [L+02] (? [R+01] suggest this applies only to North American Cretaceous forms); nearly equal heights of main cusps [L+02] [S95]; laterally compressed [C+98]; cusps have convex, roughly equal sides [S95]; height of c close to height of a [L+02]; embayment or groove on b for interlock between lower molars [L+02] lower molars with lingual cingulid reduced or absent [L+02]; mesiolabial cingular cuspule f absent [L+02]; lower molars interlock, with d/hypoconulid fitting into groove or embayment in cusp b of next more distal molar [L+02]; cusp d overhangs posterior root [S95] (contra [C+98], applies only to Jurassic forms).
Notes: [1] Rose et al. note that the opposite trend is observed in contemporary North American spalacotheriids.
Links: Mesozoic Mammals; Docodonta and Triconodonta, an internet directory; Digimorph - Arundelconodon hottoni (fossil triconodont); The Journal of Vertebrate Paleontology; ?????? Japanese).
References: Cifelli & Madsden (1998) [CM98]; Cifelli et al. (1998) [C+98]; Kermack (1963) [K63]; Luo et al. 2002) [L+02]; Rose et al. 2001) [R+01]; Sigogneau-Russell 1995) [S95]. APW 040806
 Priacodon: P. ferox Marsh 1887; P. fruitaensis Rasmussen & Callison, 1981; P. grandaevus Simpson, 1925; P. lulli Simpson, 1925; P. robustus Marsh 1879.
Priacodon: P. ferox Marsh 1887; P. fruitaensis Rasmussen & Callison, 1981; P. grandaevus Simpson, 1925; P. lulli Simpson, 1925; P. robustus Marsh 1879.
Range: Late Jurassic Kimmeridgian) to Early Cretaceous (Berriasian) of North America (WY, CO, etc) & Europe Portugal).
Phylogeny: Triconodontidae: Trioracodon + *.
Characters: cochlear housing elongate & cylindrical [J+02]; cochlea short & uncoiled [J+02]; distinct crista parotica present [J+02]; braincase bulges at level of parietal region [J+02]; bone secondary palate extends posterior to tooth row [J+02]; post-dentary trough absent [J+02]; Meckelian fossa weakly developed [J+02]; dentary angular process weak or absent [J+02]; coronoid bone absent [J+02]; splenial absent; [J+02]; dentary with pterygoid shelf, reaching dentary condyle via low crest [J+02]; coronoid process inclined at 135° [J+02]; masseteric fossa deeply excavated [RC81]; dentary condyle mediolaterally narrow, but deep & long [L+02]; dentary condyle low & faces posteriorly [RC81]; 3+ lower incisors [J+02] (contra [RC81], 2 lower incisors); i1 not enlarged or procumbent [J+02] (contra? [RC81], incisors angled 40°); large, single-rooted upper & lower canines [J+02] [RC81]; post-canine diastema present [J+02] [RC81]; premolars 4/4 [J+02] (contra [L+02] & [CM98], 3 lower premolars) (contra [RC81], premolars 3/3); premolars increase in size along tooth row [C+99] [RC81]; last P with metastylar lobe reduced or absent [J+02]; premolars without any derived characters except cusp c as large as a [J+02]; p2 a cusp unusually tall & recurved [RC81]; lower premolar cingula weakly developed [RC81]; 4-5 molars [J+02] (? [RC81], 4/4 molars with 4th reduced); no molar replacement [J+02]; m1 & M1 laterally compressed [J+02]; upper molars appear bi-lobed in lingual view [CM98]; distinct labial cingulum, but without cuspules [CM98] [J+02]; distinct lingual cingulum [CM98]; no grinding protocone [J+02]; B > C [J+02]; cusp D weakly developed [CM98]; M3 unreduced & tricuspid [RC81]; M4 bicuspid (?!) [RC81]; lower molar cusps in longitudinal row [J+02]; 4 lower molars [CM98]; lingual & labial faces roughly symmetrical [CM98]; cusps b& c of about equal height [J+02]; distinct distal cingulid [J+02]; cuspules e & f absent [J+02]; cusp d present as cuspule [J+02] [C+99]; cusp d overhangs distal root [C+99]; lower molars interlock: distal cingular cuspule d fits into groove of b cusp in next distal molar (without involvement of other cingular cuspules) [J+02]; talonid absent [J+02]; m1 labial cingulum reduced [RC81]; m4 strongly reduced [RC81]; occipital condyles cylindrical [L+02, 2]; deltopectoral crest extends past midpoint of humeral shaft [J+02]; humerus ulnar trochlea present posteriorly, with vestigial condyle anteriorly [J+02]; humeral ent- & ectepicondyle weak [J+02];
Note: [1] Luo, Kielan-Jaworowska & Cifelli, in several papers, have argued that one key feature separating Australosphenids from the "real" tribosphenics is a cingulid shelf wrapping around the mesiolingual corner of the lower molars to continue on the "lingual side of the trigonid below the paraconid." [J+02]. This feature is not present in Gobiconodon. Of course, it may not exist as a distinctive character for any taxon, as Rich argues. Alternatively, it may be meaningless in a beastie without a real trigonid.
Links: Mesozoic Mammals; Docodonta and Triconodonta, an internet directory; 416816a-s1.pdf.
References: Cifelli & Madsden (1998) [CM98]; Cifelli et al. (1999) [C+99]; Ji et al. (2002) [J+02]; Luo et al. (2002) [L+02]; Rasmussen & Callison (1981) [RC81]. APW 040806
Trioracodon: Simpson 1928, T. bisculus Marsh 1880, T. ferox Owen 1871, T. major, T. oweni.
Range: Late Jurassic Kimmeridgian) to Early Cretaceous (Berriasian) of North America (Wyoming) & Europe (England)
Phylogeny: Triconodontidae: Priacodon + *.
Characters: braincase bulges at level of parietals [L+02, 1]; petrosal (see image above at essay) very similar to Morganucodon [K63]; coronoid absent; dentary coronoid process inclined at 135° [L+02]; dentary condyle mediolaterally narrow, but deep & long [L+02]; 4 lower premolars [L+02] [CM98]; premolars increase in size along tooth row [C+99]; upper molars appear bi-lobed in lingual view [CM98]; upper molars with distinct lingual and labial cingula [CM98]; cusp D weakly developed [CM98]; only 3 lower molars [L+02] [CM98]; lingual & labial faces roughly symmetrical [CM98]; a & c cusps of about equal height; cusp d small & overhangs distal root [C+99]; mesiolingual cingulid cuspule e polymorphic [L+02].
Notes: bisculus and ferox were originally referred to Tinodon, a symmetrodont genus.
Links: Mesozoic Mammals; Docodonta and Triconodonta, an internet directory; Reconstruction of the petrosal in Late Cretaceous multituberculates (Mammalia); The Journal of Vertebrate Paleontology; The first Jurassic mammal from South America; PHYLOGENETIC INTEGRITY AND RELATIONSHIPS OF TRICONODONTIDAE ( ....
References: Cifelli & Madsden (1998) [CM98]; Cifelli et al. 1999) [C+99]; Kermack (1963) [K63]; Luo et al. 2002) [L+02]. APW 040806.
checked ATW050812