
| Cynodontia | ||
| The Vertebrates | Basal Cynodonts |
| Vertebrates Home | Vertebrate | Vertebrate |
Abbreviated Dendrogram
Therapsida
│
└─Cynodontia
├─Dvinia
└─┬─Procynosuchidae
└─┬─Galesauridae
└─Eucynodontia
├─Cynognathia
│ ├─Cynognathidae
│ └─Tritylodontidae
└─Probainognathia
├─Tritheledontidae
└─Mammaliaformes |
Contents
Index |
Cynodont evolution, after Thomas Kemp (with Wikipedia life reconstructions), copied from Mammal-like Reptiles (Korean)
 Cynodontia:
Increasingly small-sized proto-mammals with higher metabolism.
Cynodontia:
Increasingly small-sized proto-mammals with higher metabolism.
Range: from the Late Permian
Phylogeny: Eutheriodontia: Therocephalia + * : Dvinia + (Procynosuchidae + Epicynodontia).
Characters: Continued reduction of cranial dermal bone number [S01]; rostral sculpturing suggests lips and mammal-like muzzle [K82]; premaxilla vomerine process present [SH98]; maxilla does not contact prefrontal [SH98]; posterior nasal expanded with reduction of frontal so that nasal contacts lacrimal & excludes prefrontal from maxilla [HB86$] [HK01$]; prefrontal contacts postorbital and excludes frontal from orbit [SH98] [HB86$] [HK01$]; postfrontal absent [SH98] [HB86$] [HK01$]; postorbital small [K82]; parietal dominates skull table [K82]; sagittal crest anterior to parietal foramen [SH98] [HB86$]; parietal extends ventrally to cover much of lateral braincase [RS01$] [HB86$]; ventral flanges of the parietal and frontal meet flanges of opisthotic (anterior lamina) & epipterygoid; frontal contacts epipterygoid [HB86$]; 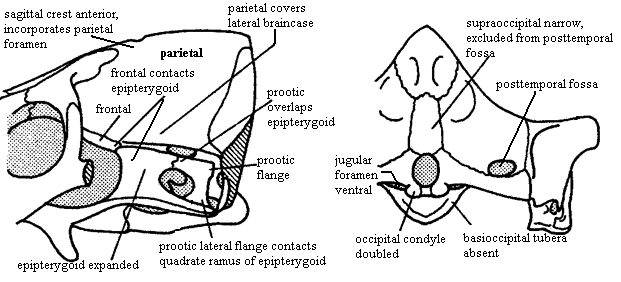 outwardly bowed zygomatic arch (suggests grinding of food by masseter muscle located behind arch) [SH98]; great anteromedial expansion of temporal fenestra [K82]; quadratojugal extends into slit in squamosal [K82]; posteroventral quadrate peg fixed in squamosal notch [SH98] [HB86$] [HK01$]; tabular surrounds posttemporal foramen [SH98] [HK01$]; supraoccipital narrow & does not contact posttemporal foramen [HB86$]; occipital condyle doubled [HB86$] [HK01$]; jugular foramen faces ventrally (not posteriorly) [HB86$]; basioccipital tubera absent [HB86$]; stapes narrowest dorsoventrally & stapedial foramen oriented dorsoventrally [SH98] [HB86$] [HK01$]; stapedial dorsal process located distally [SH98]; prootic with lateral flange [HK01$]; prootic lateral flange contacting epipterygoid [SH98]; prootic flanges overlying (vascular?) grooves [HB86$]; anterodorsal prootic overlaps posterior epipterygoid (=alisphenoid) [SH98] [HB86$]; epipterygoid expanded anteroposteriorly to proportions of mammal-type alisphenoid [RS01$] [HB86$] (now broad sheet of bone sutured to prootic & forming part of braincase wall -- not narrow rod reaching skull roof); single, enclosed trigeminal foramen [SH98]; trigeminal foramen between prootic & epipterygoid [HK01$]; vomers fused at midline [S01] [HB86$] (contra [K82] vomers paired in Dvinia); epipterygoid contacts frontal [$RS01] [SH01] [HK01$]; $? at least partially ossified secondary palate [RS01]; maxilla & palatine flange participate in secondary palate [SH98] [HB86$] [HK01$]; pterygoid transverse flanges reduced [HB86]; pterygoid quadrate ramus connected to prootic by lateral flange of prootic [K82] [HB86$] [4]; ectopterygoid does not contact maxilla [HK01$]; large dentary
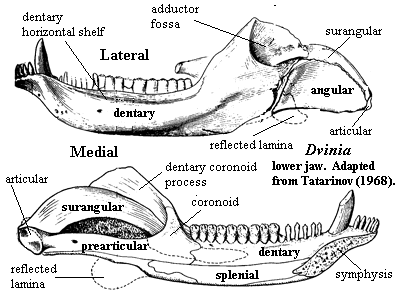
outwardly bowed zygomatic arch (suggests grinding of food by masseter muscle located behind arch) [SH98]; great anteromedial expansion of temporal fenestra [K82]; quadratojugal extends into slit in squamosal [K82]; posteroventral quadrate peg fixed in squamosal notch [SH98] [HB86$] [HK01$]; tabular surrounds posttemporal foramen [SH98] [HK01$]; supraoccipital narrow & does not contact posttemporal foramen [HB86$]; occipital condyle doubled [HB86$] [HK01$]; jugular foramen faces ventrally (not posteriorly) [HB86$]; basioccipital tubera absent [HB86$]; stapes narrowest dorsoventrally & stapedial foramen oriented dorsoventrally [SH98] [HB86$] [HK01$]; stapedial dorsal process located distally [SH98]; prootic with lateral flange [HK01$]; prootic lateral flange contacting epipterygoid [SH98]; prootic flanges overlying (vascular?) grooves [HB86$]; anterodorsal prootic overlaps posterior epipterygoid (=alisphenoid) [SH98] [HB86$]; epipterygoid expanded anteroposteriorly to proportions of mammal-type alisphenoid [RS01$] [HB86$] (now broad sheet of bone sutured to prootic & forming part of braincase wall -- not narrow rod reaching skull roof); single, enclosed trigeminal foramen [SH98]; trigeminal foramen between prootic & epipterygoid [HK01$]; vomers fused at midline [S01] [HB86$] (contra [K82] vomers paired in Dvinia); epipterygoid contacts frontal [$RS01] [SH01] [HK01$]; $? at least partially ossified secondary palate [RS01]; maxilla & palatine flange participate in secondary palate [SH98] [HB86$] [HK01$]; pterygoid transverse flanges reduced [HB86]; pterygoid quadrate ramus connected to prootic by lateral flange of prootic [K82] [HB86$] [4]; ectopterygoid does not contact maxilla [HK01$]; large dentary 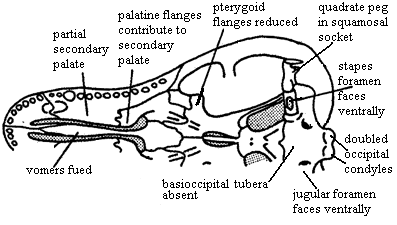 and reduced postdentary jaw bones; very large coronoid process [K82]; coronoid process with at least some lateral excavation (i.e. masseteric fossa) [RS01$] [HB86$]; posterior dentary extensively overlaps surangular [HB86$] [HK01$]; increased use of post-dentary bones for hearing; ventral surangular & anterodorsal angular emarginated to form gap between them [HB86$]; intramandibular fenestra between surangular and prearticular [K82]; angular reflected lamina small [HB86$] [SH98]; angular reflected lamina with dorsal notch closer to dentary than to articular [SH98]; articular facets of articular face posteriorly [K82]; tooth row extends under orbit [SH98]; 6 upper incisors [SH98]; incisors not serrated [SH98]; incisors spatulate [HB86$]; incisors with "smoothly ridged" cutting margins [HK01$]; canines not serrated [SH98] [HK01$]; postcanines have 2+ cusps in a line [HK01$]; postcanine teeth have distal & mesial accessory cusps [RS01] [HB86$]; lingual cingulum present [RS01] [HB86$]; lower postcanines with mesial cingulum or mesial accessory cusp [HK01$]; pterygoid dentition absent [HB86$]; atlas centrum reduced and fused to axis [K82] [SH98]; atlas pleurocentrum with broad anterior projection [SH98]; differentiation of thoracic & lumbar regions [H91]; heads of ribs in at least lumbar vertebrae are confluent [K82]; lumbar ribs reduced or absent development of diaphragm to defeat Carrier's constraint) [K82]; lumbar ribs sutured or fused to vertebrae [K82] [H91]; 4+ sacral ribs [SH98]; interclavicles with broad, parallel-sided posterior process [SH98]; scapula has laterally bent anterior and posterior edges enclosing a long vertical trough, the spinatus fossa [H91] [SH98]; coracoid and procoracoid reduced; orientation of humeral head mostly dorsal [SH98]; manus 4 with 4 phalanges [SH98]; ilium with large anterior process [K82]; ilium anterior process moderately elongated, with length
50-150% of acetabulum diameter [H91] [SH98] ([HK01] state 100-150%); pubis small and extends only just anterior to acetabulum [K82] [H91];
and reduced postdentary jaw bones; very large coronoid process [K82]; coronoid process with at least some lateral excavation (i.e. masseteric fossa) [RS01$] [HB86$]; posterior dentary extensively overlaps surangular [HB86$] [HK01$]; increased use of post-dentary bones for hearing; ventral surangular & anterodorsal angular emarginated to form gap between them [HB86$]; intramandibular fenestra between surangular and prearticular [K82]; angular reflected lamina small [HB86$] [SH98]; angular reflected lamina with dorsal notch closer to dentary than to articular [SH98]; articular facets of articular face posteriorly [K82]; tooth row extends under orbit [SH98]; 6 upper incisors [SH98]; incisors not serrated [SH98]; incisors spatulate [HB86$]; incisors with "smoothly ridged" cutting margins [HK01$]; canines not serrated [SH98] [HK01$]; postcanines have 2+ cusps in a line [HK01$]; postcanine teeth have distal & mesial accessory cusps [RS01] [HB86$]; lingual cingulum present [RS01] [HB86$]; lower postcanines with mesial cingulum or mesial accessory cusp [HK01$]; pterygoid dentition absent [HB86$]; atlas centrum reduced and fused to axis [K82] [SH98]; atlas pleurocentrum with broad anterior projection [SH98]; differentiation of thoracic & lumbar regions [H91]; heads of ribs in at least lumbar vertebrae are confluent [K82]; lumbar ribs reduced or absent development of diaphragm to defeat Carrier's constraint) [K82]; lumbar ribs sutured or fused to vertebrae [K82] [H91]; 4+ sacral ribs [SH98]; interclavicles with broad, parallel-sided posterior process [SH98]; scapula has laterally bent anterior and posterior edges enclosing a long vertical trough, the spinatus fossa [H91] [SH98]; coracoid and procoracoid reduced; orientation of humeral head mostly dorsal [SH98]; manus 4 with 4 phalanges [SH98]; ilium with large anterior process [K82]; ilium anterior process moderately elongated, with length
50-150% of acetabulum diameter [H91] [SH98] ([HK01] state 100-150%); pubis small and extends only just anterior to acetabulum [K82] [H91]; 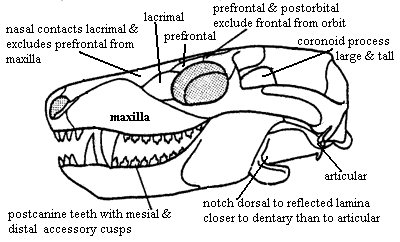 obturator foramen very large & located on puboischiadic suture [SH98]; calcaneal heel present [H91]; fore-and-aft movement of hind limb; primitively insectivores, with later carnivorous and herbivorous derived forms; some degree of endothermy likely [RS01].
obturator foramen very large & located on puboischiadic suture [SH98]; calcaneal heel present [H91]; fore-and-aft movement of hind limb; primitively insectivores, with later carnivorous and herbivorous derived forms; some degree of endothermy likely [RS01].
Notes: [1] [RS01] characterize cynodonts as showing a "fundamental reorganization of the jaw-closing musculature," with the beginning of the development of the muscular sling permitting palinal chewing. [2] from Biology 356: changes in frontal- parietal- epipterygoid structure strengthened the braincase and allowed jaw adductor muscles to have more room for their origin from the braincase. [3] from Biology 356: The excavation of the coronoid is the first indication of the mammalian masseter muscle. This was correlated with other changes in the jaw musculature. The jaw musculature of early tetrapods was composed of three muscles separated by maxillary and mandibular branches (V2 and V3) of the trigeminal nerve; the adductor mandibulae posterior, the adductor mandibulae externus, and the adductor mandibulae internus (including the pseudotemporalis and the pterygoideus). This pattern was retained in early therapsids. However, in cynodonts, most of the jaw musculature is composed of the adductor mandibulae externus. The medial portion of the externus is called the temporalis, while its new lateral portion is called the masseter (and has its origin on the zygomatic arch). [4] [HB86] refer to the quadrate ramus of the "epipterygoid." I assume this is a typographical error, something that seems to be unreasonably common in the cynodont literature.
Images: Dvinia prima courtesy of mathematical.com. Cynodont apomorphies all adapted from [HB86].
Links: Schnellbestimmung anhand von typischen Schädelmerkmalen; Prehistoric Animals; The cerebral hemispheres...; BBC Online - Walking with Dinosaurs - Fact Files; dinosaurs- dvinia prima; Reptile-Mammal very nice page); mammalogy1.html Best on the Web).
References: Hopson 1991) [H91]; Hopson & Barghusen (1986) [HB86]; Hopson & Kitching (2001) [HK01]; Kemp (1982) [K82]; Rubidge & Sidor (2001) [RS01]; Sidor (2001) [S01]; Sidor & Hopson (1998) [SH98]. ATW030209.
 Dvinia: D. prima Amalitskii (= Amilitsky)1922 (= Permocynodon sushkini).
Dvinia: D. prima Amalitskii (= Amilitsky)1922 (= Permocynodon sushkini).
Range: Late Permian of Russia, Upper Tatarian deposits of Arkhangel'sk.
Phylogeny: Cynodontia : (Procynosuchidae + Epicynodontia) + *.
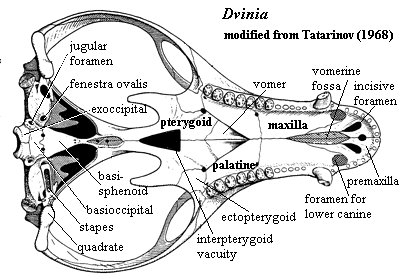
Characters: snout narrow [BS00]; distinct maxillary pit anterior to orbit [T68]; maxilla with 2 longitudinal grooves on ventral surface posterior to canines which may be attachments of labial muscles [T68]; relatively few other maxillary foramina [T68] [3]; very long, tall sagittal crest in which frontals participate [T68] [BS00]; parietal foramen absent [BS00] [T68]; sub- & postorbital bars & zygomatic arch narrow [BS00]; temporal fenestra very large, even for a cynodont [K82]; sagittal crest very high & long, without pineal foramen [BS00$] [HB86$]; sagittal crest extends posterior to occiput [K82]; squamosal lightly built [BS00]; squamosal without posterior expansion at rear of temporal fossa [HB86] [H91]; quadrate & quadratojugal exposed (not covered by squamosal) [BS00]; jaw articulation below level of occipital condyle [T68]; accessory quadratojugal- surangular articulation [T68]; posttemporal foramina relatively large [HB86] [H91]; paroccipital process anteroposteriorly narrow and almost entirely behind fenestra ovalis [HB86] [H91]; exoccipitals strongly developed [T68]; occipital condyles paired, but low and very close together, so that basioccipital is exposed only ventrally [T68]; stapes is thin-walled tube with very large stapedial foramen [T68]; stapes proximally in fenestra ovalis, distally with spur at the end of the paroccipital process [4][T68]; possible external trace of auditory meatus present on squamosal [T68]; prootic and opisthotic completely fused [T68]; prootic & pterygoid sutured above trigeminal foramen [BS00] (but contact is very restricted [HB86] [H91]); well-developed secondary palate [T68]; secondary palate 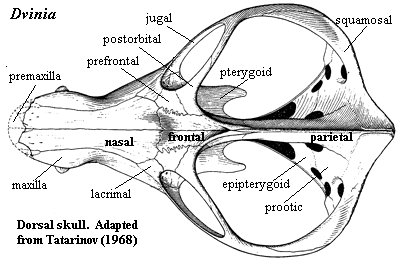 complete [BS00] (contra [K82] anterior cleft exposing vomer); foramina for lower canines in palate [T68]; vomers paired [K82] (contra [HB86] paired vomers "could not be verified"); palatal processes of maxilla & palatine meet at midline [HB86$] (contra [H91]); interpterygoid vacuity present [T68] [BS00]; ectopterygoid substantial [T68]; basipterygoid processes closely appressed & immobile [T68]; dentary small [BS00] [K82]; shelf under incisors and 3 mental foramina at root of lower canine possibly for lip musculature and circulation [T68]; weak coronoid process [BS00]; masseteric fossa restricted to dorsal portion of coronoid [K82]; postdentary bones well-developed [T68] [K82]; reflected lamina fairly large [K82]; tooth rows roughly parallel [K82]; 6 incisors [BS00] [K82]; 6 lower incisors [SH98] [HB86$]; 1 very small vestigial upper precanine [BS00] [K82]; postcanines differentiated into "premolars" and "molars" [T68] [2]; premolars slender and simple [K82]; upper postcanines bit against outer shelf of dentary [K82]; upper molars inset from margin of maxilla [SH98]; upper molars with large central cusp with 7-9 accessory cusps around margin on a cingulum [T68] [K82]; lower molars transversely expanded, with low main cusp and many labial accessory cusps [BS00$] [K82] [HB86$]; lower molars bit against secondary palate, presumably on a keratinous sheath as in dicynodonts [K82]; lower molars with main cusp displaced buccaly and ~14 marginal accessory cusps [K82]; lower molars with additional accessory cusps lingual to main cusp in central area [T68] [K82]; (lower?) lingual cingulum greatly expanded [SH98]; believed to be at least partially omnivorous, including plants and insects in diet [K82].
complete [BS00] (contra [K82] anterior cleft exposing vomer); foramina for lower canines in palate [T68]; vomers paired [K82] (contra [HB86] paired vomers "could not be verified"); palatal processes of maxilla & palatine meet at midline [HB86$] (contra [H91]); interpterygoid vacuity present [T68] [BS00]; ectopterygoid substantial [T68]; basipterygoid processes closely appressed & immobile [T68]; dentary small [BS00] [K82]; shelf under incisors and 3 mental foramina at root of lower canine possibly for lip musculature and circulation [T68]; weak coronoid process [BS00]; masseteric fossa restricted to dorsal portion of coronoid [K82]; postdentary bones well-developed [T68] [K82]; reflected lamina fairly large [K82]; tooth rows roughly parallel [K82]; 6 incisors [BS00] [K82]; 6 lower incisors [SH98] [HB86$]; 1 very small vestigial upper precanine [BS00] [K82]; postcanines differentiated into "premolars" and "molars" [T68] [2]; premolars slender and simple [K82]; upper postcanines bit against outer shelf of dentary [K82]; upper molars inset from margin of maxilla [SH98]; upper molars with large central cusp with 7-9 accessory cusps around margin on a cingulum [T68] [K82]; lower molars transversely expanded, with low main cusp and many labial accessory cusps [BS00$] [K82] [HB86$]; lower molars bit against secondary palate, presumably on a keratinous sheath as in dicynodonts [K82]; lower molars with main cusp displaced buccaly and ~14 marginal accessory cusps [K82]; lower molars with additional accessory cusps lingual to main cusp in central area [T68] [K82]; (lower?) lingual cingulum greatly expanded [SH98]; believed to be at least partially omnivorous, including plants and insects in diet [K82].
 Notes: [1] There are indications which suggest, at least to us, that Dvinia may have had the misfortune to become involved in some internal Soviet political squabble. Verbum sap. ... [2] The quotation marks are Tatarinov's. For simplicity, we will omit them in other places. [3] Tatarinov [T68] suggests that lips may have been present, but notes the general absence of the expected channels for nerves and vessels along the margin of the maxilla. However, the anterior nasals are "penetrated by numerous apertures." [4] Tatarinov [T68] speculates that the stapes was continued in cartilage to the quadrate. He includes an extensive discussion of hearing in Dvinia and the future middle ear ossicles. We have omitted this, as the results are inconclusive.
Notes: [1] There are indications which suggest, at least to us, that Dvinia may have had the misfortune to become involved in some internal Soviet political squabble. Verbum sap. ... [2] The quotation marks are Tatarinov's. For simplicity, we will omit them in other places. [3] Tatarinov [T68] suggests that lips may have been present, but notes the general absence of the expected channels for nerves and vessels along the margin of the maxilla. However, the anterior nasals are "penetrated by numerous apertures." [4] Tatarinov [T68] speculates that the stapes was continued in cartilage to the quadrate. He includes an extensive discussion of hearing in Dvinia and the future middle ear ossicles. We have omitted this, as the results are inconclusive.
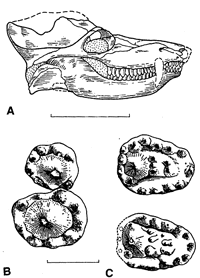
Image: (upper right) Dvinia prima, skull and right postcanines. Lingual is to the right, mesial (anterior) to the bottom. (B) upper 12th & 13th; (C) lower 11th & 13th. A strikingly derived design, but quite different from the mammalian system and vaguely reminiscent of Haramiyida. From [BS00].
Links: dinosaurs- dvinia prima; Biology 356; Chapter Two – Problems Carved in Stone.
References: Battail & Surkov (2000) [BS00]; Hopson (1991) [H91]; Hopson & Barghusen (1986) [HB86]; Kemp (1982) [K82]; Sidor & Hopson (1998) [SH98]; Tatarinov (1968) [T68]. ATW020530.
checked ATW041118, last modified MAK120206