|
|
Cretaceous |
| Mesozoic Era |
Early Cretaceous I
"Neocomian" |
The First Part of the Early Cretaceous Epoch of the Mesozoic Era: 146 to 125 million years ago
 The Early Cretaceous, as defined by the International Commission on Startigraphy, is 46 my long -- simply too long an interval to conveniently deal with as a single epoch. Our original solution on this site was to use an older system which divided the Cretaceous into Neocomian, Gallic and Senonian Epochs. However, as we are now trying our best to conform to ICS nomenclature, this is inconvenient. The Gallic includes parts of both the Early and Late Cretaceous, and tends to throw off the whole system. Accordingly, this page is now devoted to an arbitrary first half of the Early Cretaceous, including the Berriasian, Valanginian, Hauterivian, and Barremian Ages. We will continue to call this the "Neocomian" for convenience. However our demi-epoch is somewhat larger than the traditional Neocomian, which does not include the Barremian. Our new and improved Neocomian is a particularly convenient division, since it leaves the Aptian-Albian interval as a distinct demi-epoch. This second half of the Early Cretaceous is, as we will see in due course, a very different and distinctive time period.
The Early Cretaceous, as defined by the International Commission on Startigraphy, is 46 my long -- simply too long an interval to conveniently deal with as a single epoch. Our original solution on this site was to use an older system which divided the Cretaceous into Neocomian, Gallic and Senonian Epochs. However, as we are now trying our best to conform to ICS nomenclature, this is inconvenient. The Gallic includes parts of both the Early and Late Cretaceous, and tends to throw off the whole system. Accordingly, this page is now devoted to an arbitrary first half of the Early Cretaceous, including the Berriasian, Valanginian, Hauterivian, and Barremian Ages. We will continue to call this the "Neocomian" for convenience. However our demi-epoch is somewhat larger than the traditional Neocomian, which does not include the Barremian. Our new and improved Neocomian is a particularly convenient division, since it leaves the Aptian-Albian interval as a distinct demi-epoch. This second half of the Early Cretaceous is, as we will see in due course, a very different and distinctive time period.
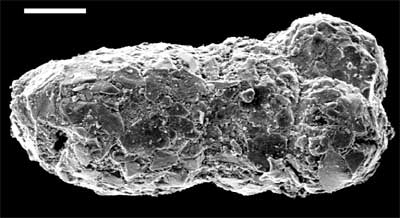
Image: The Barremian brought the beginning of the great Cretaceous flowering of the Foraminifera. The image shows Bulbobaculites, one of these new forams. The scale bar is 100µ. The image is from the extensive catalog of images and information compiled by David Haig at Barremian - Turonian Foraminifera from the Southern Carnarvon Platform. This appears to be a part of an even larger site, the BioStrat Gallery, which houses a number of such collections. ATW040811.
For perhaps 300 million years, terrestrial animal types had been global in extent, there being only a single supercontinent, Pangea, across which creatures could freely roam. Now, for the first time, a seperation of continents had produced global geographic isolation, and provincialism and a unique faunas appeared on different landmasses. This, the last period of dinosaurian evolution, was also the period of their greatest diversity, although only a fraction of the types (mainly latest Cretaceous Western North American and Asian forms) are known.
It was only during the early Cretaceous period that the supercontinent of Pangea began to break up. This is indicated by map where we see the continents finally beginning to separate and take on their present forms. As C.B. Cox explains:
"A spreading axis gradually extended clockwise round Africa until it was almost completely isolated, and India began its long northward journey....Shallow seas separated Asia from Europe, and penetrated far towards the centre of North America.....The gap between North...and South America may have widened somewhat..., and any archipelagic connection between the Americas may have become rather more...difficult as a dispersal route for terrestrial animals." [p.76]
As the centrifugal process of continental fragmentation gets underway, the continents of Gondwana began to break and separate. And as there was a process of diversification of the land-masses, so there was also of diversification of animal-life; both in numbers of different kinds and in their geographical isolation. For the first time, real provincialism appears. Of interest is the fact that India has started to break away from the rest of Gondwanaland, to become an island continent all on its own. India in fact was to become the Australia of the Mesozoic. MAK
|
Harland Epoch |
ICS Epoch
|
Age
|
ICS
Base (mya)
|
Duration
|
Gallic
|
|
Cenomanian |
99.6 |
6.1 |
|
|
Albian |
112 |
12.4 |
| Aptian |
125.0 |
13.0 |
|
|
Barremian |
130.0 |
5.0 |
Neocomian
|
Hauterivian |
136.4 |
6.4 |
| Valanginian |
140.2 |
3.8 |
| Berriasian |
145.5 |
5.3 |
Late Jurassic
|
|
Tithonian |
150.8 |
5.3 |
ICS (2004). Geowhen Database
ATW040811.
 We are accustomed to hearing that the climate of the Mesozoic was hot and humid -- like August in Houston, Texas, but lasting about 200 million years. Those who find this boring and oppressive will be relieved to know that the climate of the Neocomian was not so predictable. In fact, temperatures dropped through the Late Jurassic to produce what was, by Mesozoic standards, an "ice-house" world during parts of the early Neocomian. This "ice house" would still seem relatively warm (and, yes, even humid) by comparison with the dry and frigid standards of our own day; but the dinosaurs found it rather brisk, none the less. The poles had small permanent ice caps, and snow probably fell on the cool temperate forests that surrounded the poles. This was the coldest portion of the entire Mesozoic.
We are accustomed to hearing that the climate of the Mesozoic was hot and humid -- like August in Houston, Texas, but lasting about 200 million years. Those who find this boring and oppressive will be relieved to know that the climate of the Neocomian was not so predictable. In fact, temperatures dropped through the Late Jurassic to produce what was, by Mesozoic standards, an "ice-house" world during parts of the early Neocomian. This "ice house" would still seem relatively warm (and, yes, even humid) by comparison with the dry and frigid standards of our own day; but the dinosaurs found it rather brisk, none the less. The poles had small permanent ice caps, and snow probably fell on the cool temperate forests that surrounded the poles. This was the coldest portion of the entire Mesozoic.
We know that things warmed up fairly rapidly after the middle of the Berriasian, but the warming trend may have faltered during the Valanginian. Our knowledge of the Valanginian climate is full of uncertainties. The indications are that the Valanginian included a sharp cold snap severe enough to reduce sea levels due to a (probably brief) build-up of polar ice. There are also paradoxical indications of a temperature spike and of deep ocean anoxia, on the one hand, and increased ocean primary productivity, on the other, during the Valanginian. These have have left the matter in doubt. Notwithstanding this brief and uncertain reversal, world temperatures climbed rapidly after the Valanginian, possibly increasing as much as 17° C. before the end of the Early Cretaceous (i.e., the better-known heat wave of the middle Aptian).
Links: There are a number of good web sites which touch on the cimate of the Neocomian. Among these are: Early Cretaceous; Early Cretaceous Climate; Climate History; BBC - Science & Nature - Dinosaurs; CLIMATE-OCEAN ISOTOPIC SIGNALS FROM LATE Jurassic-EARLY Cretaceous ...; Ocean Drilling Program- Leg 198 Preliminary Report; OZ fossils - The Age of Reptiles - The Climate. ATW040811, revised ATW041102.
Neocomian Life
 The Neocomian is best known for the first appearance of flowering plants. During Neocomian time itself, the angiosperms probably did not amount to much, as the image suggests. Nevertheless, they became a detectable part of the flora and would go on to dominate the world's vegetation by the end of the Cretaceous. The development of flowers led immediately to their co-evolution with insects. So we begin to see specialized pollinators evolving from several insect clades.
The Neocomian is best known for the first appearance of flowering plants. During Neocomian time itself, the angiosperms probably did not amount to much, as the image suggests. Nevertheless, they became a detectable part of the flora and would go on to dominate the world's vegetation by the end of the Cretaceous. The development of flowers led immediately to their co-evolution with insects. So we begin to see specialized pollinators evolving from several insect clades.
Even more important -- to us, at least -- the Neocomian was probably the time in which therian mammals evolved and diversified into placental and marsupial clades, and no doubt other groups which have left no Cenozoic descendants. Birds likewise became common in this period, as did cerapod dinosaurs -- the hypsilophodonts, heterodontosaurs, and, by the end of the Neocomian, iguanodonts.
The Neocomian may have been the high point of the pterosaurs, with such large and specialized types as Ornithocheirus, and of the Mesozoic breeds of turtles, which produced some enormous forms. It was the last period in which ichthyosaurs were at all common in the oceans.
ATW050216. Text public domain. No rights reserved.
 During the Neocomian, rudist reefs began to dominate over more conventional reefs formed by scleractinian corals. Coral reefs were the predominant form in the Jurassic, just as they are today. Rudists "reefs" are not quite reefs, and the rudists themselves are certainly not corals. Rudists are actually a group of aberrant bivalve mollusks which became extinct at the end of the Cretaceous. More about rudist reefs in general may be found in our discussion of Aptian Reefs. Many sources state that rudist reefs didn't become important until the Aptian-Albian, but this depends on where you look. The Central Tethyan region -- roughly between Arabia in the southeast to France in the Northwest -- was Rudist Country as far back as the Berriasian. However, areas such as the Caribbean Basin were dominated by corals until the Aptian.
During the Neocomian, rudist reefs began to dominate over more conventional reefs formed by scleractinian corals. Coral reefs were the predominant form in the Jurassic, just as they are today. Rudists "reefs" are not quite reefs, and the rudists themselves are certainly not corals. Rudists are actually a group of aberrant bivalve mollusks which became extinct at the end of the Cretaceous. More about rudist reefs in general may be found in our discussion of Aptian Reefs. Many sources state that rudist reefs didn't become important until the Aptian-Albian, but this depends on where you look. The Central Tethyan region -- roughly between Arabia in the southeast to France in the Northwest -- was Rudist Country as far back as the Berriasian. However, areas such as the Caribbean Basin were dominated by corals until the Aptian.
The difference may be related to mode and speed of growth. We don't know much about the speed of rudist growth as such. However, rudists probably made quick, sloppy structures. They trapped sediment between "stalks," to form a "reef" out of sand and silt. Corals do it the hard way. A coral reef is built, entirely of calcareous tests, grown and deposited cell by cell, over millenia. The entire reef is biogenic. This is slow work. We may imagine, then, rudists moving in like weeds whenever something happened to disrupt the standing crop of coral. They would quickly overgrow the old framework, covering everything over with sand, silt, and fragments of shell and thus preventing the old, complex, high-diversity communities from re-establishing themselves -- a bit like suburban housing developments, in fact.
Our guess about rudist development is reasonably well substantiated by geology. The Tethys was notably unstable during the Neocomian, as Europe was cracked and splintered by the growing Tethys Seaway. In addition to geological activity, the region also seems to have been pounded by frequent, severe storms. Finally, the region experienced three periods of rapid sea level increase: (1) early Valanginian to early Hauterivian (this episode may have occurred in two stages), (2) middle Hauterivian, and (3) late Hauterivian to early Barremian. During these periods, the carbonate slopes on which coral reefs had developed "drowned." That is, the sea rose and the platforms came to be submerged below the zone optimal for photosynthesis. Because of the high relief of the sea bottom (e.g., abrupt cliffs caused by faulting) the corals couldn't grow into shallower water.
 This was a sort of worst case for reef stability, and this is one of the areas to which rudists spread earliest in the Cretaceous. In fact, within the Tethyan realm, conditions were a little worse yet toward the eastern end, in present-day eastern Europe. Conditions comparable to the later mid-Cretaceous black shales are particularly well known in this region from well within the Neocomian. Not surprisingly, this seems to have been the original home of the rudists. They are known in this sector from the Late Jurassic, and they became the dominant reef-builders at the very beginning of the Neocomian.
This was a sort of worst case for reef stability, and this is one of the areas to which rudists spread earliest in the Cretaceous. In fact, within the Tethyan realm, conditions were a little worse yet toward the eastern end, in present-day eastern Europe. Conditions comparable to the later mid-Cretaceous black shales are particularly well known in this region from well within the Neocomian. Not surprisingly, this seems to have been the original home of the rudists. They are known in this sector from the Late Jurassic, and they became the dominant reef-builders at the very beginning of the Neocomian.
On the other side of the proto-Atlantic, in the future Gulf of Mexico, we see a very different pattern. A great barrier reef stretched from Texas to the southern tip of Florida. The northern Gulf of Mexico was sheltered from the ocean on all sides. No violent tectonic forces were active beyond a gentle tug extensional stretching) as the Yucatan attempted to turn in slow circles like a cat preparing for a serious nap. Sea levels also rose here, but the topography was so flat that growth on the northern margin easily kept pace with drowning to the south. This is practically ideal reef territory, and we see no rudists at all until the Aptian, when temperature extremes may have stressed the coral communities beyond endurance. A similar pattern is seen on the coast of what is now Nova Scotia. As we might expect, complex, intermediate patterns are observed in France, Jamaica, and Venezuela where rudists established themselves in the Hauterivian or Barremian.
In short, the rudists seem to be a classic case of evolution by adventitious specialization. They are not more "fit" than corals. Indeed, they eventually lost out to the corals. However, they could grow quickly and thus move in whenever the coral reefs suffered a setback. They then established themselves to the exclusion of the former owners. A good deal of "competitive replacement" in evolution must actually work this way
-- not by head-to-head competition, but by slipping in the back door while no one is home.
Image credit: reef image from Sedimentology MO8313 course materials, Oxford Brookes University (lots of good stuff on reefs of all kinds at this site).
ATW041026. Text and map public domain. No rights reserved.
 The dinosaurian megafauna of the Neocomian was very different to that of the Late Jurassic. Gone are the diverse cetiosaurid, omeisaurid, and diplodocid sauropods, the range of spined and plated stegosaurs, and the ornitholestid, ceratosaurid, torvosaurid, and allosaurid guilds that fed upon them. In their place are titanosaurs, Iguanodonts (left), polacanthid ankylosaurs, primitive ceratopsians, the strange semi-aquatic spinosaurs, and an astonishing range of small and large coelurosaurs, including a number of new clades (Ornithomimosaurs, Dromaeasaurs, Troodonts, Therizinosaurs, Tyrannosaurs) that clearly seem to have arisen from the poorly known maniraptors of the late Jurassic, and of course a huge radiation of birds, taking the place of small and medium-sized pterosaurs. Other dinosaurs continued with relatively little change - brachiosaurs and dicraeasaurs among the sauropods and hypsilophodonts among the ornithopods - while others again, like the stegosaurs, were greatly reduced in number.
The dinosaurian megafauna of the Neocomian was very different to that of the Late Jurassic. Gone are the diverse cetiosaurid, omeisaurid, and diplodocid sauropods, the range of spined and plated stegosaurs, and the ornitholestid, ceratosaurid, torvosaurid, and allosaurid guilds that fed upon them. In their place are titanosaurs, Iguanodonts (left), polacanthid ankylosaurs, primitive ceratopsians, the strange semi-aquatic spinosaurs, and an astonishing range of small and large coelurosaurs, including a number of new clades (Ornithomimosaurs, Dromaeasaurs, Troodonts, Therizinosaurs, Tyrannosaurs) that clearly seem to have arisen from the poorly known maniraptors of the late Jurassic, and of course a huge radiation of birds, taking the place of small and medium-sized pterosaurs. Other dinosaurs continued with relatively little change - brachiosaurs and dicraeasaurs among the sauropods and hypsilophodonts among the ornithopods - while others again, like the stegosaurs, were greatly reduced in number.
In his classic work The Dinosaur Heresies, Dr Bob Bakker suggests that the replacement (in Laurasia) of Sauropods by Ornithischia (iguanodonts and ankylosaurs) was brought about through angiosperm evolution, itself triggered by selection pressures from dinosaurian feeding, with the slow growing conifers that the sauropods fed upon being replaced by fast-growing and fast regenerating angiosperms, which in turn became food for the new low-browsing ornithischian dinosaurs. But this hypothesis, persuasive and fascinating as it seems, ignores the fact that titanosaurian sauropods continued as the predominate dinosaur clade in Gondwana right until the end of the Mesozoic, and that Angiosperms themselves did not become an important part of the floral community until the Campanian (near the end of the Cretaceous). Nevertheless, it is quite likely that selection pressures and feedback loops between dinosaurs and the vegetation they fed upon may have been an important contributing factor in the evolution of Mesozoic plants and of course of the dinosaurs themselves.
In the late Jurassic, with the exception of China (presumably geographically isolated) with its unique omeisaur-stegosaur megafauna, dinosaurs were very similar from one continent to the next, differeing only at the species and generic level. But in the Cretaceous the break up of Pangea and the resulting island continents led to the start of a growing provincialism, in contrast to the uniformity of the Jurassic world. Psittacosaurs and therizinosaurs flourished in Asia, and hypsilophodonts, polacanthids, and brachiosaurs diversified in Europe and North America. Other clades like iguanodonts and titanosaurs seem to have been universal. As the Cretaceous progressed, the various continents would each become home to a distinctive fauna.
Iguanodon at a waterhole - illustration © BBC/ABC from Walking with Dinosaurs
MAK050104.
page uploaded on Kheper Site on 28 May 1998, page uploaded on Palaeos Site 9 April 2002,
last modified ATW040811, MAK050104.
checked ATW040810

