{kind=link}
{kind=link}
 The Basal Osteolepiforms
The Basal Osteolepiforms| Sarcopterygii | ||
| The Vertebrates | Osteolepiformes (2) |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Teleostomi
├─Neopterygii
└─Sarcopterygii
├─┬─Onychodontiformes
│ └─Actinistia
└─Rhipidistia
├─Dipnomorpha
└─┬─Rhizodontiformes
└─Osteolepiformes
├─Osteolepididae
└─┬─Megalichthyidae
└─┬─Canowindridae
└─┬─Tristichopteridae
└─Elpistostegalia
├─Panderichthys
└─Tetrapoda
|
Contents
Overview |
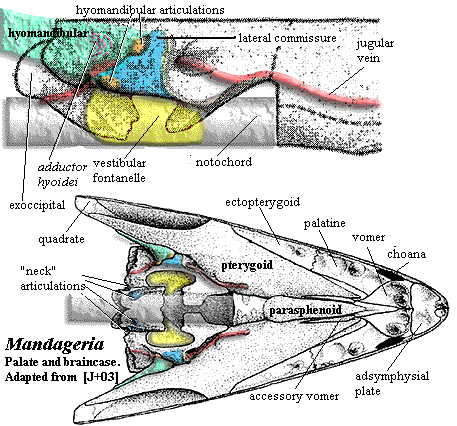
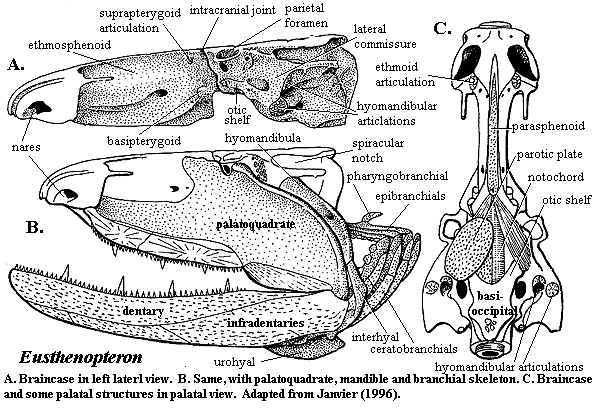
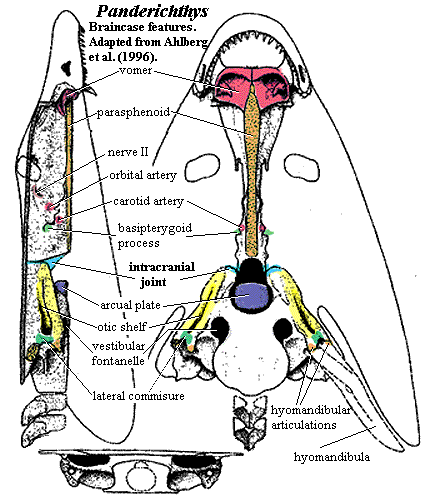
We have discussed the dermal skull of tetrapodomorphs. On the whole, it makes pretty good sense from a mechanical point of view. Developments inside the skull are somewhat less tidy, as they often are. The structure of the osteolepiform braincase, represented by Eusthenopteron has been described earlier. Since we lack good information for the rhizodonts, this can tentatively be taken as primitive for the tetrapodomorphs. This obviously represents an inadequate sample, since Eusthenopteron is itself a tristichopterid, not a basal "osteolepid" at all. However, recent descriptions of the braincases of an osteolepid and a megalichthyid are said to be entirely consistent with this model. Johanson et al. (2003). The only other braincases known with reasonable certainty in this area of phylospace come from the derived tristichopterid Mandageria and the elpistostegalian Panderichthys. As always, we must make do with what brains we have.
The attentive reader (although if you had listened in class you would probably not be reading this site) may wish to refer to the following images in addition to the image of Mandageria shown here:
a) braincase of Eusthenopteron
b) braincase of Panderichthys.
In all of these forms, the anterior braincase is relatively narrow. In Mandageria, and probably Cabonnichthys, the parasphenoid plate is wider. The vomers have lateral processes and/or accessory vomerine plates which clasp the sides of the parasphenoid. This is consistent with what we see in the dermal skull, as the midline support in the dermal skull is being reflected in a strong median support for the palate through the parasphenoid. However, this is not the pattern in the elpistostegalians. Here, the parasphenoid has only narrow palatal exposure, accessory vomerine plates are lacking, and the posterior process of the vomer is slight.
The hint of an explanation may come from the nasal capsules. With one exception, the nasal capsules of all osteolepiforms are believed to be very similar to those of Eusthenopteron. They are large and fully ossified. The exception is Panderichthys. Here, the nasal capsules are smaller and, at least in the view of Vorobyeva & Schultze (1991), unossified posteriorly. In even the most primitive tetrapods, the nasal capsule is never ossified at all. Clack 2002).
What may be going on here is another basic divergence in design. As we have discussed, one of the common themes of tetrapodomorph evolution is the lengthening of the rostrum. The problem with a long rostrum is not only its internal weakness, but the torque it can generate on the skull; and, the further out on the rostrum the weight is placed, the more awkward it becomes. Structures like big vomers and bony nasal capsules are very expensive in terms of torque, because they occur at the very distal end of the head. There are two ways to deal with an outlying structure which is difficult to support and generates torque: add more braces, or reduce the distal weight of the structure. Tristichopterids seem to have emphasized the former method, and elpistostegalians the latter.
This makes for an interesting, if speculative, point. Water is a very supportive medium. Torque can be a problem for a moving aquatic creature which needs to change direction quickly. The animal may need to apply torque to move the rostrum. But there are other strategies to overcome this problem [1]. Torque is of far larger concern to a creature on land. Even sitting still, a terrestrial animal must constantly fight the torque created by gravity. Thus, there are many long-snouted animals in the oceans, but few on land. We may then suppose that elpistostegalians were either subject to selective pressure from terrestrial living, or were singularly pre-adapted to adopt that mode of life.
Another peculiarity is found in the middle of the braincase, at the intracranial joint. Some tristichopterids show a tendency to obliterate the joint externally. Elpistostegalians seem to have eliminated completely this flexible gap between the parietals and postparietals. However, no tetrapodomorph appears to have made the corresponding changes in the braincase. According to Ahlberg et al. (1996), the separation between anterior and posterior braincase is as distinct in Panderichthys as it is in Eusthenopteron. These workers argue that some relatively sudden and dramatic change in braincase structure must have occurred with the advent of tetrapods, since no tetrapod has a divided brain.
Posteriorly, both osteolepiform lineages show some tendency to close up the basicranial opening and to move the vestibular fontanelles forward. Perhaps this is another area in which some fairly rapid and important changes occurred at the transition from tetrapodomorph to tetrapod. The interesting example of Mandageria shows that this was possible, although it sheds no immediate light on the tetrapod condition. The ventral vacuities in this tristichopterid were rearranged to create the functional equivalent of a flexible neck, although the structure bears no similarity to the tetrapod cervical apparatus.
In short, the braincase was, at the same time, conservative and idiosyncratic. The braincase does not show the same kind of long-term, lineage-independent concerted change we observe in the dermal skull. This overall conservatism does not seem to be the result of some tight functional constraint. To the contrary, it appears that the braincase was capable of rapid and significant evolutionary change, a potential realized in Mandageria and -- quite differently -- in tetrapods. If the braincase was capable of responding to selective pressure but showed no consistent pattern of change, we can reach only one conclusion: that it was simply not subject to any consistent external selective forces. ATW031031.
GOTO PART 4. The Palate, Jaw, & Branchial Skeleton
[1] For example, changing the shape of the rostrum to generate lift in the desired direction. A long extension actually helps, if the shape change can be made to the more distal regions.
The Basal OsteolepiformsWe do not cover the basal osteolepiform families in much detail. The basal "osteolepiform" morphotype has already been taken up in detail elsewhere. These families, Osteolepididae, Megalichthyidae and Canowindridae, differ relatively little from the morphotype.
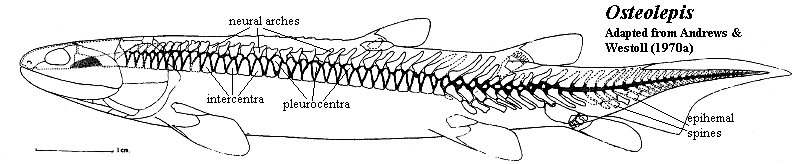
The Osteolepididae we take to be a small monophyletic group which might be defined as Osteolepis > Eusthenopteron. The family includes Osteolepis, Gogonasus, probably Thursius, and perhaps a few others. The Osteolepidids differ from other osteolepiforms in having epicercal tails and rounded fins. Generally, osteolepiforms have homocercal, or even trilobate, tail fins. The second dorsal and the anal fin are well-separated from the caudal fin. The vertebral centra are relatively well ossified throughout the vertebral column, but are not fused into rings. Rather, they have a typical rachitomous arrangement, with two large intercentra surrounding the notochord ventrally and laterally. The intercentra fuse ventrally except in the mid-trunk region. The pleurocentra are small, always separate, and apparently articulate only with the base of the neural arches anterior to them. The pleurocentra bear unique, "pulley" structures dorsally, consisting of a smooth central channel flanked by two "knobs." See image below. The scapulocoracoid is weakly attached to the cleithrum, and the humerus is relatively flat and featureless. As with all of the basal families, the scales are rhomboid and covered with cosmine.
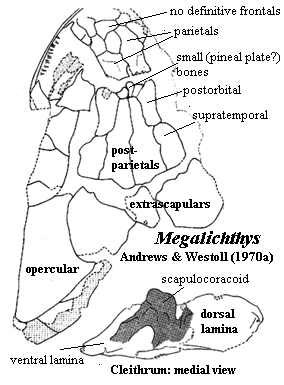
The Megalichthyidae are a more distinct group. The skull is primitive. The nares are relatively dorsal and defined by specialized tectals. The intracranial joint is strongly marked on the dermal skull, with a series of small dermal bones inserted between the parietal and postparietal. Frontals are completely absent. A single, median anterior "fang" is present, but it is carried on a palatal process of the premaxilla. Since it lacks a replacement pit, it is not a true fang. The vomers do not articulate with the parasphenoid (as is also the case with osteolepidids). The vertebrae show considerable variation, but are generally holospondylous and ring-like, with no trace of a separate pleurocentrum. As in osteolepidids, the posterior fins are well separated from the tail fin. However, megalichthyids are unique in having the anal fin articulate with the spine at a position well posterior to the articulation of the second dorsal. Externally, the tail fin is almost symmetrical. As in osteolepidids, the scapulocoracoid is neither massive nor strongly secured to the cleithrum, which has a small anterior process and weak attachment to the clavicle. The cosmine coat is a bit less extensive than in osteolepidids.
The Canowindridae were a diverse group of basal osteolepiforms with a wide distribution in the Late Devonian. The family includes at least Beelarongia, Canowindra, Koharolepis, and probably Gyroptychius. They show some derived features probably acquired independently from other groups. Thus, for example, the postorbital is excluded from the orbit, and the jugal and postorbital are short, but very large. The skull is flat, and some derived genera have rounded scales with an internal boss, lacking cosmine. These are all features found in other clades, notably the Tristichopteridae. However, canowindrids lack the postspiracular bone and other synapomorphies of the tristichopterids and are clearly a distinct, if closely-related group. ATW031101.
Image: (of fossil) Osteolepis from the University of Birmingham, Lapworth Museum.
 Osteolepididae: Gogonasus, Osteolepis, Thursius?. Probably paraphyletic [CA96]. We have tried to restrict this taxon to a minimal, monophyletic group.
Osteolepididae: Gogonasus, Osteolepis, Thursius?. Probably paraphyletic [CA96]. We have tried to restrict this taxon to a minimal, monophyletic group.
Range: Middle Devonian (Eifellian) to Guadalupian (Sakmarian) [CA96]. Possibly as early as Early Devonian (Emsian) [J80].
Phylogeny: Osteolepiformes: (Megalichthyidae + (Canowindridae + (Tristichopteridae + Elpistostegalia))) + *.
Characters: generally small (<50 cm) [L95]; small bones cover rostrum & may be superficially fused [J96]; lateral tectal contacts supraorbital [J96]; postorbitals & jugal short [J96]; squamosal large [J96]; anterior braincase short [J80]; snout broad & blunt [J96]; nasal capsules well-separated & completely ossified [J80]; vomers meet anterior to, & do not overlap, parasphenoid [JA97]; parasphenoid broad & denticulated [J96]; parasymphysial fang pair usually absent [JA97]; parasymphysial plate usually large, contacting coronoid [JA97]; inner surfaces of centra elements probably roughened for notochord contact as in Eusthenopteron [AW70a]; centra more extensively ossified than in Eusthenopteron [C88]; intercentra paired in trunk, but fused anteriorly and caudally [C88]; intercentra continuous with hemal arch in tail [C88]; intercentra in half-rings, fused ventrally except in mid-trunk [AW70a]; intercentra generally uniform throughout column [AW70a]; pleurocentra very large, especially anteriorly, & fused dorsally except in mid-trunk [AW70a] (contra [C88]: pleurocentra always paired); pleurocentra only associated with base of next anterior neural arch [AW70a]; pleurocentra with "pulley" structure dorsally (smooth central channel flanked by two "knobs") [AW70a]; pleurocentra bear small hemispherical pits which receive neural arches both anteriorly and posteriorly [AW70a]; in caudal region, 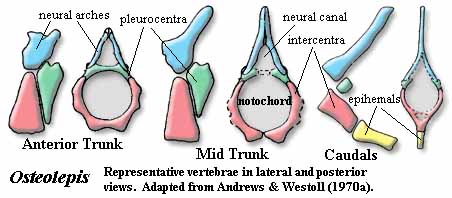 pleurocentra reduced to small, irregular bones not closely integrated with intercentra [AW70a]; neural arches massive, but short (especially anteriorly), fused dorsally in caudal series [AW70a]; neural arches smooth externally & probably surrounded a dorsal ligament just above the neural canal [AW70a]; parapophyses and ossified ribs absent [AW70a]; fins rounded [J96]; two dorsal fins [L95]; anal fin basal without rod-like proximal section [AW70a]; anal fin basal articulates with 2 hemals which fit into "steps" (rectangular indentations) in the basal [AW70a]; anal fin with 3-4 distal radials [AW70a]; caudal fin heterocercal/epicercal [L95] [J96]; caudal fin lobes probably supported by neural spines and jointed epihemals [AW70a]; posterior part of cleithrum cosmine covered, while anterior is ornamented with parallel vertical ridges [J80]; tripodal scapulocoracoid [J80]; humerus broad and flat compared with Eusthenopteron [JA97]; rhomboid, cosmine-covered scales [J96]; cosmine frequently on dermal bones [J96]; cosmine frequently absent from cleithrum [AW70a]; scapulocoracoid not firmly attached to cleithrum [AW70a].
pleurocentra reduced to small, irregular bones not closely integrated with intercentra [AW70a]; neural arches massive, but short (especially anteriorly), fused dorsally in caudal series [AW70a]; neural arches smooth externally & probably surrounded a dorsal ligament just above the neural canal [AW70a]; parapophyses and ossified ribs absent [AW70a]; fins rounded [J96]; two dorsal fins [L95]; anal fin basal without rod-like proximal section [AW70a]; anal fin basal articulates with 2 hemals which fit into "steps" (rectangular indentations) in the basal [AW70a]; anal fin with 3-4 distal radials [AW70a]; caudal fin heterocercal/epicercal [L95] [J96]; caudal fin lobes probably supported by neural spines and jointed epihemals [AW70a]; posterior part of cleithrum cosmine covered, while anterior is ornamented with parallel vertical ridges [J80]; tripodal scapulocoracoid [J80]; humerus broad and flat compared with Eusthenopteron [JA97]; rhomboid, cosmine-covered scales [J96]; cosmine frequently on dermal bones [J96]; cosmine frequently absent from cleithrum [AW70a]; scapulocoracoid not firmly attached to cleithrum [AW70a].
Links: Osteolepiforms and the ancestry of tetrapods abstract, arguing paraphyly); Lebedev another abstract, also espousing paraphyly); Osteolepis macrolepidotus image of fossil); Fishes several large images, but specimen quality is variable); CROSSOPTERYGII as always on this site, generously large images of very high-quality fossils); Ammoniten; www.fossilien.de || Quastenflosser aus Schottland ||; Urweltmuseum Neiderhell - Sonderausstellung "Der Fisch, der an ...; Devonian Times - unidentified megalichthyid; Urweltmuseum Neiderhell - Sonderausstellung "Der Fisch, der an ....
References: Andrews & Westoll (1970a) [AW70a], Carroll 1988) [C88], Cloutier & Ahlberg (1996) [CA96], Janvier (1980) [J80], Janvier (1996) [J96]; Johanson & Ahlberg (1997) [JA97], Long (1995) [L95]. ATW031018.
Megalichthyidae
: Cladarosymblema, Ectosteorhachis, Mahalalepis, Megalichthys, Megapomus, Megistolepis.Range: Mississippian to Pennsylvanian Guadalupian!? [L95])
Phylogeny: Osteolepiformes:: Canowindridae + (Tristichopteridae + Elpistostegalia)) + *.
Characters: to 1m [L95]; specialized tectals surrounding nares [L95]; nares somewhat dorsally placed [J96]; frontals absent [AW70a] [1]; braincase fissure reflected on dermal skull [AW70a] [1]; series of small bones between parietals & postparietals [AW70a] [1]; large anterior folded teeth (fangs?) on a posterior process of the premaxilla [J80]; vomers wide [J96]; lateral extrascapulars approach closely on midline [JA97]; premaxilla with long posterior palatal process [L95] premaxilla palatal process with large tooth dividing anterior palatal vacuity, but not sloping down into vacuity [JA97]; medial tooth behind unbroken premaxillary tooth row [JA97]; vomers may meet anterior to parasphenoid without overlapping it [JA97]; vomers lack posterior process [JA97]; parasymphysial fang pair present in some [JA97]; vertebrae show considerable individual variation [AW70a]; holospondylous, with pleurocentra absent or completely fused [AW70a]; ribs absent [AW70a]; no canal for longitudinal ligament (as contrasted with Osteolepis) [AW70a]; neural arch extends posteriorly from anterodorsal corners of centrum; caudal vertebrae small compared to most osteolepiforms [AW70a] [3]; caudal hemal and neural spines thin, long and sharply pointed [AW70a]; 2nd dorsal & anal fin supports morphologically similar to Eusthenopteron [AW70a]; 2nd dorsal and anal fins articulate with only one truncated spine (with two in various other forms) [AW70a]; anal fin articulates 3-4 vertebrae behind 2nd dorsal fin (compare, 1 in Eusthenopteron) [AW70a]; anal fin with 3 distal radials [AW70a]; caudal fin lobes supported by neural spines and jointed epihemals [AW70a]; epichordal lobe enlarged [AW70a]; tail approaches external symmetry [AW70a]; cleithrum with broad, oblong dorsal lamina & ventrally tapering, triangular ventral lamina [AW70a]; cleithrum anterior process small or absent [AW70a]; cleithrum, bulge over scapulocoracoid small [AW70a]; cleithrum external surface with cosmine limited, sometimes absent (much variation among specimens), and always with an unornamented, cosmine-free anterior crescent, presumed to be for attachment of membrane lining back of gill chamber [AW70a]; cleithrum with very narrow external clavicular overlap ventral to membrane attachment area [AW70a]; cleithrum medial surface smooth, without roughened attachment areas for scapulocoracoid [AW70a]; clavicle with strong posterior process articulating with medial face of cleithrum [AW70a]; clavicle dorsal process unusually long, forming double articulation with cleithrum as in Rhizodus [AW70a] [4]; clavicle external surface ornamented with tubercles and a few ridges, rarely with cosmine [AW70a] clavicle also with anterior smooth area for membrane attachment [AW70a]; scapulocoracoid tripodal, very similar to Eusthenopteron,but "very lightly [and superficially] ossified [AW70a]; scapulocoracoid somewhat dorsoventrally elongate [AW70a]; scapulocoracoid glenoid foramen [2] conspicuous [AW70a]; glenoid fossa as in Eusthenopteron, or possibly wider in comparison to length [AW70a]; fin with large basal scutes [AW70a]; 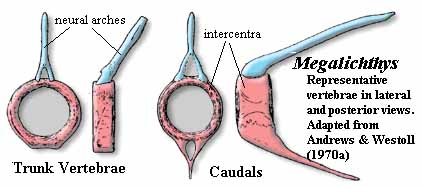 pectoral fin lepidotrichia covered with scales, and leading edge with hemicylindrical ridge scales, both sets covered in cosmine [AW70a]; lepidotrichia in short segments [AW70a]; pectoral fin skeleton poorly known, but appears to be similar to Eusthenopteron, but with bones thinner and more elongate [AW70a]; scales with cosmine [L95].
pectoral fin lepidotrichia covered with scales, and leading edge with hemicylindrical ridge scales, both sets covered in cosmine [AW70a]; lepidotrichia in short segments [AW70a]; pectoral fin skeleton poorly known, but appears to be similar to Eusthenopteron, but with bones thinner and more elongate [AW70a]; scales with cosmine [L95].
Notes: [1] these points are evident from the figures in [AW70a], but not commented on by those authors. [2] this is not the glenoid fossa itself, but a large foramen located above the dorsal margin of the glenoid, at one end of the supraglenoid buttress. Unfortunately, the glenoid is turned away from the viewer in the image, which shows the cleithrum from an anteromedial view. [3] This pattern resembles those of the Dipnoi. Note the marked difference between this structure and that of Osteolepis. Thursius and Gyroptichius show somewhat intermediate structures. Eusthenopteron has vertebrae with an even looser pattern than Osteolepis, but with a short rib stabilizing the connection between intercentra and neural arches. [AW70a]. [4] In Megalichthys, the dorsal spine of the clavicle passes medial to the cleithrum, while the ventral overlap is lateral. This is reversed in Rhizodus. [AW70a].
Links: Comparaciones anatómicas; Megalichthys hibberti; 247.htm; Teeth.
References: Andrews & Westoll (1970a) [AW70a], Janvier 1980) [J80], Janvier (1996) [J96], Johanson & Ahlberg (1997) [JA97], Long 1995) [L95]. ATW031018.

Canowindridae: Beelarongia, Canowindra, Gyroptychius?, Koharolepis.
Range: Middle Devonian? to Late Devonian (Famennian) [CA96] of Antarctica [L95], Australia, Greenland, Europe
Characters: skull wide & flat [L95]; eyes very small [L95]; marked intracranial joint; broad postparietal shield [CA96]; lateral extrascapulars triangular & approach closely on midline [CA96] [JA97]; postorbital excluded from orbit [CA96] [JA97]; jugal & postorbitals short [J96]; jugal extremely large [JA97]; operculars large & deep [L95]; primitively with rhombic, cosmine-covered scales, lost in more derived forms and replaced by rounded scales with medial boss, as in Tristichopteridae [L95].
Phylogeny: Osteolepiformes::: (Tristichopteridae + Elpistostegalia) + *.
 Image: Gyroptychius upper) from Quastenflosser: Einige Ausstellungsobjekte der Sonderausstellung; Canowindra grossi lower) adapted from Vickers-Rich & Rich (1999).
Image: Gyroptychius upper) from Quastenflosser: Einige Ausstellungsobjekte der Sonderausstellung; Canowindra grossi lower) adapted from Vickers-Rich & Rich (1999).
Links: Canowindra grossi Age of Fishes Museum); 242.htm; 241.htm; 340.htm Gamagori Museum site); Paleozoic Fossils UK; Urweltmuseum Neiderhell - Sonderausstellung "Der Fisch, der an ...; Geologisch Museum - Genesis in Artis; Fishes.
References: Cloutier & Ahlberg (1996) [CA96], Janvier 1996) [J96]; Johanson & Ahlberg (1997) [JA97], Long 1995) [L95]; Vickers-Rich & Rich (1999). ATW031005.
checked ATW021227